 2014, Vol.41
2014, Vol.41


构建STK11基因融合质粒检测其对肺癌细胞迁移能力的影响
引用本文
张鑫君,邱祥南,章龙珍,王建设,丁 昕,辛 勇,覃朝晖,姚元虎. 构建STK11基因融合质粒检测其对肺癌细胞迁移能力的影响[J]. 肿瘤防治研究, 2014, 41(07): 707-710. 
ZHANG Xinjun,QIU Xiangnan,ZHANG Longzhen,WANG Jianshe,DING Xin,XIN Yong,QIN Zhaohui,YAO YuanHu. Construction of STK11 Gene Fusion Plasmid and Its Effect on Migration of Lung Cancer Cells. Cancer Research on Prevention and Treatment, 2014, 41(07): 707-710.
构建STK11基因融合质粒检测其对肺癌细胞迁移能力的影响
221002 江苏徐州,徐州医学院附属医院放射治疗科 徐州医学院肿瘤生物治疗实验室
摘要:目的 构建表达STK11(serine/threonine kinase 11,STK11)基因和报告基因EGFP融合蛋 白的重组质粒,并转染肺癌A549和H460细胞株,检测其对肺癌细胞迁移能力的影响。方法 构建重 组质粒pEGFP-STK11,双酶切和测序进行鉴定。重组质粒转染A549和H460细胞株,荧光显微镜观察 EGFP蛋白表达情况,Western blot检测STK11蛋白表达情况,划痕实验检测其对肺癌细胞迁移能力的 影响。结果 酶切和测序结果表明pEGFP-STK11重组质粒构建成功;在转染A549和H460细胞后24h观 察到绿色荧光最强;Western blot结果显示STK11蛋白在转染后24 h条带最深;划痕实验显示,转染组 细胞迁移距离小于0.9%氯化钠溶液对照组(P<0.05)。结论 成功构建STK11基因重组质粒,并能转 染A549和H460细胞,转染后对肺癌细胞迁移能力起一定抑制作用。
关键词:
STK11
细胞迁移
A549
H460
Construction of STK11 Gene Fusion Plasmid and Its Effect on Migration of Lung Cancer Cells
Department of Radiation Oncology, Affiliated Hospital of Xuzhou Medical College, Xuzhou Medical College Tumor Biologial Therapy Institute, Xuzhou 221002, China
Abstract:Objective To construct eukaryotic expression plasmids containing enhanced green fluorescent protein(EGFP) gene and serine/threonine kinase 11(STKK11) gene, and to explore its effect on migration of A549 and H460 cells. Methods Fusion plasmid of pEGFP-STK11 was constructed and identified by double restriction enzyme digestion and gene sequencing. The resultant plasmid was transfected into A549 and H460 cells respectively. The expression of EGFP and STK11 were observed by fluorescence microscopy and Western blot analysis. The cell migration ability was tested by Wound-healing assay. Results Enzyme digestion and DNA sequence analysis showed the recombinant vector of pEGFP-STK11 was constructed successfully. Fluorescence microscopy and Western blot analysis indicated STK11 gene was overexpressed in A549 and H460 cells after transfection, and reached the peak after 24 h. Wound-healing assay revealed cell migration distance of STK11 gene transfected group was significant less than that of control group (P<0.05). Conclusion STK11 gene fusion plasmid is constructed and transfected into lung cancer cell lines A549 and H460 successfully. Cells migration is inhibited to some extent after transfection.
Key words:
Serine/threonine kinase 11(STK11)
Cells migration
A549
H460
0 引言
2.2 EGFP 表达和STK11基因蛋白表达
2.3 划痕实验检测细胞迁移能力
3 讨论
STK11(serine/threonine kinase 11,STK11) 基因最早是从Peutz-Jeghers 综合征(Peutz-Jeghers syndrome,PJS)患者中发现的,目前被认为是一 种抑癌基因,参与多个生物学过程和信号通路[1]。 我们构建含目的基因STK11的EGFP融合载体转染 A549和H460细胞,并探讨其对肺癌细胞迁移能力 的影响。 1 材料与方法 1.1 材料
健康者胚肾细胞系293和人肺癌细胞株A549、 H460细胞、EGFP-N1、大肠埃希菌DH5α由徐州医学院肿瘤生物治疗实验室保存;人H460细胞购 买于南京凯基生物有限公司;引物合成及测序工 作由上海生工生物工程技术服务有限公司完成; RPMI 1640培养液购自赛默飞世尔生物化学有限公 司;胎牛血清购于浙江天杭生物科技公司;限制 性内切酶、连接酶、RT-PCRKit Ver.2试剂盒、凝 胶回收试剂盒、质粒中量提取试剂盒均购自北京 TIANGEN公司;脂质体(lipofectamine 2000) 购于 Invitrogen 公司;STK11抗体、辣根过氧化物酶标 记的二抗购于abcam公司。 1.2 引物设计及STK11基因扩增
根据GenBank的LKB1基因序列(N M 000455.4),利用Primer predict和DNAclub软件 设计引物,引物由上海英俊生物技术有限公司合 成。引物上下游分别引入Hin dⅢ/KpnⅠ酶切位 点。上游引物:5’-ATAGCTAGCATGGAGGTGGT GGACCCG-3’,下游引物:5’-AGAGAATTCCTG CTGCTTGCAGGCC-3’。
按照Trizol试剂盒说明书,从293细胞中提取总 RNA,获得STK11全长。 1.3 重组质粒构建及鉴定
将末端加A后的目的基因STK11连接到PMD18- T载体上,得到pMD18- T- STK11重组 质粒。分别双酶切质粒pEGFP-N1和pMD18-TSTK11,获得pEGFP-STK11重组质粒。 用含阳性克隆的细菌抽提质粒 ,对质粒p EGF P - S TK11进行NheⅠ和EcoRⅠ双酶切, 37℃、2h,1%琼脂糖凝胶电泳鉴定。重组质粒送 交上海英俊生物技术有限公司进行基因测序,将 测序结果与质粒中插入序列的ORF对比,验证重 组质粒插入片段的正确性。 1.4 转染及实验分组
转染前一天,将处于对数生长期的细胞用 0.25%胰酶消化,分别将A549和H460细胞浓度调 整至为(2~5)×107 cells/ml,接种至6孔板,待细 胞铺满板底80%~90%时,按照Lipofectamine 2000 转染试剂说明书进行转染。实验分pEGFP-STK11 重组质粒转染组和0.9%氯化钠溶液对照组。每组6 个复孔。 1.5 荧光显微镜观测和Western blot检测
在转染后12、24、48和72 h荧光显微镜下分 别观测两组细胞EGFP的表达情况。分别在转染后 24、48和72 h收集细胞,按照细胞蛋白提取试剂盒 要求提取蛋白,−80℃冰箱保存,Western blot检测 STK11蛋白表达。 1.6 划痕实验
在6孔板背面画平行直线做标记,每孔3条。 分别将两组细胞常规消化后,5×105细胞/孔种植到 6孔板内过夜,待细胞基本融合达到80%时,弃去 培养液,用PBS清洗三遍后,用10 μl微量移液头 在6孔板内垂直于标记纵向划痕,PBS冲洗3次去 除细胞,加入无血清培养液继续培养。倒置显微 镜下,在3个不同的划痕部位拍照,48 h后在同样 的位置再拍照观察创伤愈合情况,应用Photoshop 软件计算划痕间距。实验重复3次,细胞迁移率= (划痕后即刻测量的划痕间距-划痕后48 h测量的 划痕间距)/划痕后即刻测量的划痕间距×100%。 1.7 统计学方法
应用SPSS19.0软件对实验数据进行统计分 析。统计描述、均数采用x±s表示,两组间均数比 较采用t检验,P<0.05为差异有统计学意义。 2 结果 2.1 重组质粒鉴定
酶切鉴定结果显示在1 300 bp左右出现条带, 与STK11基因大小相同,见图 1。对pEGFP-STK11 插入片段进行测序,结果显示插入序列无突变, 与GenBank中STK11序列一致。
 |
图1 pEGFP-STK11重组质粒双酶切鉴定结果 Figure 1 Double restriction enzyme digestion identif ication result of pEGFP-STK11 recombinant plasmid |
A549细胞和H460细胞在转染后各时间点,荧 光显微镜下均可见pEGFP-STK11重组质粒组细胞 呈绿色,细胞形态完好,在转染后24 h EGFP表达 最强,见图 2;0.9%氯化钠溶液对照组细胞在转染 后各时间点,荧光显微镜下均未观察到绿色荧光。 在转染后各时间点,Western blot检测结果均 显示与目的蛋白分子量一致的目的条带,24 h时条 带颜色最深;而0.9%氯化钠溶液对照组未见目的 条带表达,见图 3。
 |
A-D:Fluorescence microscopy detection of SKT11 gene expression after A549 cells transfected with pEGFP-STK11 for 12,24,48 and 72h, respectively;E-H:Fluorescence microscopy detection of SKT11 gene expression after H460 cells transfected with pEGFP-STK11 for 12,24, 48 and 72 h, respectively
图2 pEGFP-STK11质粒转染A549和H460细胞后绿色荧光表达情况( ×100)
Figure 2 Fluorescent microscopy detection of A549 and H460 cells transfected with pEGFP-STK11 ( ×100) |
 |
Control:0.9% NaCl control group; 24 h:cells transfected with pEGFPSTK11 for 24 h;48 h:cells transfected with pEGFP-STK11 for 48 h;72 h: cells transfected with pEGFP-STK11 for 72 h
图3 Western blot检测STK11蛋白表达
Figure 3 STK11 expression detected by Western blot |
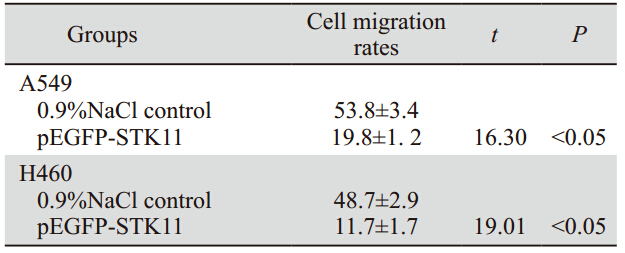
与划痕后即刻测量相比,各组细胞在划痕后48 h,划痕距离均明显缩小,有融合的趋势,见图 4。两种细胞均显示pEGFP-STK1重组质粒转染组 划痕较0.9%氯化钠溶液对照组细胞愈合慢,转染 组细胞迁移率小于0.9%氯化钠溶液对照组,差异 有统计学意义(P<0.05),且H460细胞迁移率小 于A549细胞,见表 1。
 |
A,B: wound closure of A549 cells in 0.9%NaCl control group after 0 and 48 h; C,D: wound closure of A549 cells in pEGFP-STK11 group after 0 and 48 h; E,F: wound closure of H460 cells in 0.9%NaCl control group after 0 and 48 h; G,H: wound closure of H460cells in pEGFP-STK11group after 0 and 48 h, respectively
图4 A549和H460细胞划痕实验检测细胞迁移情况 ( ×200)
Figure 4 Wound closure of A549 and H460 cells detected by Wound-healing assay( ×200) |
|
|
表 1 A549和H460细胞划痕实验结果(x±s, %, n=3) Table 1 Migration rates of A549 and H460 cells detected by Wound-healing assay(x±s, %, n=3) |
STK11基因位于人类染色体19p13.3区域上, DNA全长2155 bp,编码区含1302 bp,包括9个外 显子,其转录的mRNA约3.0 kb[2] 。STK11编码的丝氨酸/苏氨酸蛋白激酶包含433个氨基酸残基, 功能结构域包含一个激酶区域和氨基端非催化区 域的核定位信号通路[3]。STK11虽然最早在PJS综 合症中发现,但是有文献报道其在非小细胞肺癌 中也有散发性突变,突变率高达30% [4]。据文献报道,我们选择的H460和A549肺癌细胞株STK11基 因存在37密码子突变,无STK11蛋白表达[5]。针对这两种细胞株进行STK11重组质粒转染,使之过表 达,更有利于研究STK11基因的生物学功能。对我 们构建的重组质粒双酶切琼脂电泳可显示1 302 bp 的目的基因条带,基因测序与GenBank中STK11序 列一致,提示STK11重组质粒成功构建。荧光显微 镜检测pEGFP-STK11重组质粒转染组细胞EGFP表 达,也提示STK11重组质粒成功构建。Western blot 结果显示转染后细胞有STK11蛋白条带出现,表明 重组质粒在细胞内正确表达。
STK11基因调节多项生物学过程包括细胞生 长、细胞周期、极化、转移和能量代谢[6]。高益 军等[7]报道STK11通过下游mTOR-HIF-1α信号通路 调控肿瘤转移相关基因LOX(lysyl oxidase)而介 导肺癌的转移,LOX上调可激活β-1-integrin信号 通路,增强细胞的侵袭能力。文献报道,在临床上观察到发生远处转移的肺癌患者中STK11突变 率增高[8]。我们的细胞划痕实验提示,重组质粒转 染组细胞有细胞划痕愈合减慢的现象,初步证实 STK11有抑制肺癌迁移作用。我们下一步会采用 Transwell等方法进一步验证STK11基因的迁移抑制 作用,并通过检测Lox、Pdgf R和Vegfc等转移基因 来探讨其可能机制。
参考文献
| [1] | van Lier MG, Wagner A, Mathus-Vliegen EM, et al. High cancer risk in Peutz-Jeghers syndrome: a systematic review and surveillance recommendations[J]. Am J Gastroenterol, 2010,105(6): 1258-64. |
| [2] | Hemminki A, Markie D, Tomlinson I, et al. A serine/threonine kinase gene defective in Peutz-Jeghers syndrome[J]. Nature, 1998,391(6663):184-7. |
| [3] | Alessi DR, Sakamoto K, Bayascas JR. Lkb1-dependent signaling pathways[J]. Annu Rev Biochem, 2006,75:137-63. |
| [4] | Zhong D, Guo L, de Aguirre I, et al. Lkb1 mutation in large cell carcinoma of the lung[J]. Lung Cancer, 2006,53(3):285-94. |
| [5] | Sun LL, Zhong DS, Wu S, et al. Establishment and gene expression profiling of LKB1 stable knockdown lung cancer cell line[J]. Chin Med J (Engl), 2011,124(13):2028-32. |
| [6] | Lützner N, De-Castro Arce J, R?sl F. Gene expression of the tumour suppressor LKB1 is mediated by Sp1, NF-Y and FOXO transcription factors[J]. PLoS One, 2012,7(3):e32590. |
| [7] | Gao Y, Xiao Q, Ma H, et al. LKB1 inhibits lung cancer progression through lysyl oxidase and extracellular matrix remodeling[J]. Proc Natl Acad Sci U S A, 2010, 107(44):18892-7. |
| [8] | Matsumoto S, Iwakawa R, Takahashi K, et al. Prevalence and specificity of LKB1 genetic alterations in lung cancers[J]. Oncogene, 2007, 26(40): 5911-8. |