 2014, Vol.41
2014, Vol.41
外源性TGFBI抑制乳腺癌细胞生长的体内外实验
引用本文
张鹤美,贺金奖,高四海,尉 红,张增利,童 建,Tom K·Hei,李冰燕. 外源性TGFBI抑制乳腺癌细胞生长的体内外实验[J]. 肿瘤防治研究, 2014, 41(07): 693-697. 
ZHANG Hemei, HE Jinjiang, GAO Sihai, WEI Hong, ZHANG Zengli, TONG Jian. TGFBI Inhibits Proliferation of Breast Cancer Cell in vitro and in vivo. Cancer Research on Prevention and Treatment, 2014, 41(07): 693-697.
外源性TGFBI抑制乳腺癌细胞生长的体内外实验
1. 215123 苏州,苏州大学公共卫生学院卫生毒理学教研室;
2. 哥伦比亚大学放射研究中心
2. 哥伦比亚大学放射研究中心
摘要:目的 研究转化生长因子β诱导基因 (transforming growth factor-β induced gene, TGFBI) 是否
能抑制人乳腺癌细胞株(MDA-MB-231)的体内外增生。方法 将外源性TGFBI稳定转染到人乳腺癌细
胞株(MDA-MB-231)中,测定细胞增殖率、细胞周期、软琼脂克隆形成率及P21、P53蛋白表达变化,
检测乳腺癌细胞的致瘤性。结果 外源性TGFBI在人乳腺癌细胞株(MDA-MB-231)中能够稳定地高表
达;外源性TGFBI可以明显地抑制乳腺癌细胞因血清生长因子刺激的增生;与转染空质粒的对照组细
胞V23101比较,外源性TGFBI相对软琼脂克隆形成数减少了90.89%; TGFBI可以使人乳腺癌细胞阻
滞在G1期,延缓其进入S期的时间。外源性TGFBI可以延长裸鼠肿瘤发生的潜伏期,并降低肿瘤发生
率。结论 TGFBI能够抑制人乳腺癌细胞株(MDA-MB-231)的体内外增生。
关键词:
转化生长因子β诱导基因
人乳腺癌
细胞增殖
体内
体外
TGFBI Inhibits Proliferation of Breast Cancer Cell in vitro and in vivo
ZHANG Hemei1, HE Jinjiang1, GAO Siai1, WEI Hong1, ZHANG Zengli1, TONG Jian1, Tom K·Hei2, LI Bingyan1

1.Department of Health Toxicology, School of Public Health, Soochow University, Suzhou
215123, China;
2. Center for Radiological Research, Columbia University Medical Center, New York, U.S.A.
2. Center for Radiological Research, Columbia University Medical Center, New York, U.S.A.
Abstract:Objective To explore if transforming growth factor-β induced gene (TGFBI) could inhibit
the proliferation of human breast cancer cell lines MDA-MB-231 in vitro and in vivo. Methods Ectopic
TGFBI was stably transfected into MDA-MB-231. Cell proliferation, cell cycle, soft agar cloning efficiency,
protein expression of P21 and P53 were analyzed to test tumorigenicity of human breast cancer cells.
Results Ectopic TGFBI was expressed highly and stably in MDA-MB-231 cells, with the significant
decrease of cells proliferation rate. Compared with V23101 transfected with empty plasmid, ectopic TGFBI
expression resulted in a significant decrease of relative soft agar colony formation number by 90.89%. Tumor
cells transfected with TGFBI were arrested in the G1 phase and delayed into the S phase. Ectopic TGFBI
reduced tumorigenicity by 16.99% and delayed the incubation of tumor growth. Conclusion TGFBI could
inhibit the proliferation of human breast cancer cell in vitro and in vivo.
Key words:
TGFBI
Human breast cancer
Proliferation
in vitro
in vivo
0 引言
2.2 外源性TGFBI对乳腺癌细胞增殖的影响
2.3 外源性TGFBI对乳腺癌细胞锚着独立性生长的影响
2.4 外源性TGFBI对乳腺癌细胞周期的影响
2.5 TGFBI对细胞周期调节蛋白P21和P53表达的影响
2.6 TGFBI对乳腺癌细胞体内致瘤性的影响
2.6.1 外源性TGFBI对裸鼠成瘤性的影响
2.6.2 外源性TGFBI对肿瘤组织细胞增殖的影响
3 讨论
乳腺癌在女性癌症死因中占首位[1],且因其新 发病例最多[2],发病年龄呈年轻化趋势,严重威胁 女性健康。但乳腺癌患病原因和恶性进程的精确分 子机制仍不十分清楚。 转化生长因子β诱导基因(transforming growth factor-β induced gene,TGFBI)也被称为Betaig-3, 是一种由TGF-β诱导的分泌蛋白,最初认为其与肿 瘤相关,是因为发现TGFBI能够使接种裸鼠体内肿 瘤细胞的致瘤能力减弱[3]。最近研究发现,TGFBI 的FAS1结构域有抑制肿瘤作用,表现在抑制肿瘤 生长、血管发生及促凋亡作用[4]。Zhao等[4,5,6,7,8]发现很 多肿瘤细胞中TGFBI表达下降,并且体内、外实验 都发现外源性TGFBI能够抑制肿瘤细胞的恶性转 变。
本研究将外源性TGFBI转染到人乳腺癌细胞 (MDA-MB-231)中,得到稳定性表达TGFBI的细胞 株,观察TGFBI对乳腺癌细胞的生长和侵袭是否具 有抑制作用。 1 材料与方法 1.1 TGFBI转染及鉴定
人乳腺癌细胞株(MDA-MB-231)来源于美国 模式培养物集库存,使用DMEM完全培养液(英杰 公司,美国),10%胎牛血清( Gibco公司,美国), 100 µg/ml 链霉素,100 u/ml青霉素。TGFBI转染 的主要过程为:制备4 µg载体pRc/CMV2-TGFBI或 pRc/CMV2-空载体与转染试剂Lipofectamine Plus (英杰公司,美国)混合后,加入已接种细胞的6孔 培养板培养3 h;然后换完全培养液继续培养72 h。按1∶10比例把已转染TGFBI或空载体的细胞 分到100 mm的培养板中,在400 mg/ml 的G418培 养液(Sigma公司,美国)中培养,挑选稳定性表达 TGFBI或空载体的抗性集落。
采用实时定量PCR检测TGFBI mRNA的表达。 TRIzol (Invitrogen公司,美国) 提取细胞总RNA; RT试剂盒(英杰公司,美国)将提取的总RNA反 转录成cDNA;实时定量 PCR仪7300 (AB公司,美 国),用SYBR Green定量分析TGFBI mRNA。内参 基因GAPDH (目录号PPH00150E) ,靶基因TGFBI (目录号PPH01904B)。PCR循环参数:95℃ 15 min;95℃ 30 s,55℃ 30 s,72℃ 30 s,40个循环。 实验结果以倍数的变化2-∆∆Ct表示,ΔCt = CtTGFBI- CtGAPDH ΔΔCt = Ct样品-Ct对照。
细胞培养24 h提取蛋白,加10%的三氯乙酸后 离心,用丙酮洗涤沉淀物两次,室温干燥;加50 µl 上样缓冲液煮沸5 min,上样,Western blot法检 测细胞培养上清液中TGFBI蛋白的表达。免疫组 织化学方法鉴定TGFBI在细胞内分布情况。使用 染色试剂盒 (Vector Laboratories公司,美国),先用 1.5%马血清封闭样品,加1:1 000的人抗TGFBI单 抗(Minneapolis公司,美国)37℃孵育2 h,PBS 洗3次后;加1:5 000的二抗,室温孵育30 min,加 VECTASTAIN Elite ABC室温反应30 min,最后加 DAB ( 3,3’ - 二氨基联苯胺)底物显色6 min,双蒸水 冲洗;酒精梯度脱水后,加封片剂封固样品。在 BX60 奥林巴斯显微镜下观察TGFBI蛋白在细胞内 的分布。 1.2 细胞增殖测定
细胞增殖测定试剂盒(英杰公司,美国)检 测细胞增殖,先用无血清的 DMEM培养液培养细 胞36h,然后换用含 10% 胎牛血清的 DMEM 培 养液培养 0、12、16、20及24后用PBS冲洗,加 入100 µl染料结合液,37℃孵育1 h。使用酶标仪 (Bio-Tec公司,美国)测定荧光强度。细胞增殖 率用增加的荧光强度百分数表示,即增加的百分 比 = ( n 时的荧光强度 - 0时的荧光强度) /0时的荧 光强度。 1.3 软琼脂克隆形成测定
将1 000个细胞与1 ml 0.35%琼脂糖混合,接 种于底部铺有0.75%琼脂糖的培养皿中,每组4个 平行样本。在10倍显微镜下观察并计算集落数, 实验结果用相对集落数表示,即相对克隆形成数 = 外源性TGFBI表达细胞的克隆形成数 /母代细胞的 克隆形成数。 1.4 细胞周期分析
无血清培养液培养细胞36 h,使细胞周期同 步化于G0/G1期。换用含10%胎牛血清的培养液培 养,在0、4、8、12、16、20及24 h后,收集包括 培养液中悬浮的全部细胞,75%冰乙醇4℃固定过 夜,用PI标记细胞,流式细胞仪(BD公司,美国)分 析细胞周期。 1.5 Western blot检测
将细胞接种于100 mm的培养皿中,使细胞 同步化于G0/G1期,换用10%胎牛血清培养液刺激 细胞进入新的细胞周期,在血清刺激后的不同时 间点,用冷的PBS冲洗细胞。用细胞裂解液(50 mMTris-Hcl,pH8,150 mM Nacl,1%SDS,1mM PMSF)提取蛋白质,用蛋白浓度试剂盒(美国伯 乐公司)测蛋白浓度,取30 µg蛋白质加入到50 µl 的上样缓冲液中煮沸5 min,上样于10%~20%SDS 胶(伯乐公司,美国),在电压20V的条件下电泳 2~4 h,在半干燥条件下将蛋白样品转移到PVDF膜 上。洗膜,5%脱脂牛奶封闭,分别加入1∶1 000 的P21、P53或β-actin一抗,4℃过夜,洗膜三次; 1∶5 000的二抗孵育2 h,洗膜三次;ECL显色液 (Amersham Biosciences公司,美国),全自动洗片 机(KODAK公司,美国)进行全自动显色、洗片 和烘干。 1.6 体内致瘤性检测
2月龄裸鼠饲养在特殊的无病原体的环境中, 小鼠可以自由进食,饮水,日光/黑暗为12 h/12 h。 每组皮下注射5×106个肿瘤细胞于裸鼠背部。每周 称体重,观察肿瘤的发生情况,注射细胞16周后 处死裸鼠。 1.7 免疫组织化学染色
将裸鼠皮下取出的肿瘤组织用10%福尔马林固 定一周左右,送病理室进行脱水、石蜡包埋、切片 后,进行Ki67和MECA-32的免疫组织化学染色。 一抗分别是Ki67和MECA-32,浓度为1∶100。每个样品随机选择5个视野共100个细胞,计数细胞 核或胞质呈棕黄色的Ki67或MECA-32的阳性细胞 数,数据以相对阳性细胞百分比表示,即阳性细 胞减少率 =( 实验组-对照组)/ 对照组。 1.8 统计学方法
采用SPSS 17.0进行统计学分析,各组实验数 据均重复3次,以均数±标准差 (x±s) 表示。先作 方差齐性检验,再用t-test进行两个均数之间的比 较。P<0.05为差异具有统计学意义。 2 结果 2.1 TGFBI稳定性表达细胞株的鉴定
将外源性pRc/CMV2-TGFBI载体转染到 MDAMB-231中,转染细胞株命名为T231,从中挑选 T23108、23109和T23113细胞株用于本研究。实 时定量PCR结果显示,T23108、23109和T23113细 胞株中TGFBI mRNA表达明显高于其母代MDAMB-231细胞,与永生化乳腺上皮细胞MCF-10F中 的表达一致,见图 1A。免疫组织化学结果显示, T23113的细胞质和细胞核中均有TGFBI蛋白的表 达,见图 1B。Western blot结果显示,T23113细胞 培养上清液中TGFBI蛋白也明显高于母代细胞, 见图 1C。以上结果表明,外源性TGFBI转染后得 到了稳定性高表达的乳腺癌细胞株,接近于永生 化乳腺上皮细胞MCF-10F中TGFBI的表达。
 |
A: the fold change of TGFBI mRNA expression level of the
stable transfected cells calculated by 2-∆∆Ct; B: TGFBI protein expression in MDA-MB-231 transfected with TGFBI detected by
immunohistochemical staining (DAB); C: TGFBI protein expression in
MDA-MB-231 transfected with TGFBI detected by Western blot
图1 外源性TGFBI稳定性转染的乳腺癌细胞株中mRNA和蛋白表达
Figure 1 Expression of mRNA and protein in breast cancer cells transfected with ectopic TGFBI |
图 2的结果表明,母代MDA-MB-231细胞在 血清刺激后12 h和24 h的增殖率分别为5.02%和 64.44%,而外源性TGFBI表达的T23108、T23109 和T23113细胞在血清刺激后的12 h,细胞几乎没有 增殖,在血清刺激后24 h的平均增殖率为39.62%。 说明外源性TGFBI可以明显地抑制乳腺癌细胞因 血清生长因子刺激的增殖。
 |
Cell proliferation was assessed by CyQUANT NF proliferation
kit at indicated times after serum stimulation. Proliferation rate was expressed as increased percentages = ( fluorescence density at nh – fluorescence density at 0h)/Fluorescence density at 0 h
图2 稳定转染外源性TGFBI与母代乳腺癌细胞株MDAMB-231增殖率比较
Figure 2 Comparison of proliferation rates of parental
ectopic MDA-MB-231 cells and cells transfected with ectopic TGFBI |
与转染空质粒的V 2 3 1 0 1细胞相比,外源 性TGFBI表达的细胞T23113,形成了较少的克 隆(P<0.01),相对软琼脂克隆形成数减少了 90.89%,表明TGFBI显著地抑制了乳腺癌细胞的 锚着独立性生长能力,见图 3。
 |
Data were expressed as relative colonies = (colonies in TGFBIexpression tumor cells) / (colonies in parental tumor cells), *: P< 0.01,
compared with V23101 transfected with vector control
图3 转染外源性TGFBI与空质粒的乳腺癌细胞株锚定独立性生长能力比较
Figure 3 Comparison of anchorage independent growth of
breast cancer cell lines transfected with ectopic TGFBI and MDA-MB-231 cells |
细胞周期同步化后,外源性TGFBI表达和母 代乳腺癌细胞处在G1期的比例分别是68.57%和 64.29%。血清刺激后的不同时间,与母代细胞相 比较,外源性TGFBI表达的细胞处于 G1期的比例 明显增多(P<0.05),而处于S期细胞比例减少, 尤其是在血清刺激后16 h更为明显(P<0.05)。表 明外源性TGFBI使乳腺癌细胞阻滞于G1期,延缓 细胞分裂的进程,结果与2.2对细胞增殖的影响结 果相一致。
 |
Line graph and plot illustrated the distribution of cells in the G1 and S
phases over a time period of 32h
图4 外源性TGFBI对乳腺癌细胞周期的影响
Figure 4 Effects of ectopic TGFBI on cell cycle of breast
cancer cells |
转染空载体的V23101细胞与母代乳腺癌细胞 一样,在血清刺激后12和24 h 时P21和P53表达没 有明显变化,而外源性TGFBI表达的T23113细胞在血清刺激12和24 h时,P21和P53表达明显增多, 见图 5。实验结果提示,TGFBI是通过上调P21和 P53表达延缓乳腺癌细胞进入S期的。
 |
图5 外源性TGFBI对乳腺癌细胞P21和P53瞬时表达的影响 Figure 5 Effects of ectopic TGFBI on temporal expressions of breast cancer cells P21 and P53 |
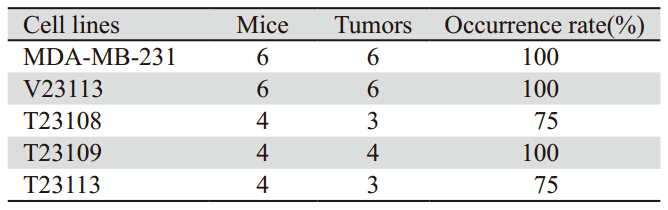
在注射母代乳腺癌细胞后第4周,裸鼠开始出 现肿瘤包块,至第5周时,所有注射母代和空载 体的MDA-MB-231细胞的裸鼠都发现肿瘤,肿瘤 发生率100%;而注射外源性TGFBI表达T23108、 T23109和T23113细胞的裸鼠至第6周时才发现有 肿瘤包块。至注射后第12周时,TGFBI组的肿瘤 发生率为83.33%[(3+4+3)/ (4+4+4)],见表 1。说明 外源性TGFBI表达使裸鼠肿瘤发生潜伏期延长了2 周,发生率也减少了16.7%。
|
|
表 1 转染TGFBI的乳腺癌细胞对裸鼠体内肿瘤发生率的影响 Table 1 Effects of breast cancer cells transfected with ectopic TGFBI on tumor occurrence rate of nude mice in vivo |
Ki67是肿瘤组织中细胞增殖的标志蛋白,可 证明TGFBI对体内肿瘤细胞生长的影响。结果显 示,与对照组相比较,注射T23108、T23109和 T23113细胞后,肿瘤组织中Ki67表达位于细胞核 内,呈棕黄色,见图 6A,阳性细胞的百分比平均 减少了33.84%,见图 6B。说明外源性TGFBI可以 抑制肿瘤组织中细胞的增殖,这与TGFBI抑制乳 腺癌细胞体外生长的结果一致。
 |
A: Immunohistochemical staining of Ki67 protein in nude mice
subcutaneously injected with breast cancer cells transfected with
TGFBI;B: data showed relative percentage of positive cells, *: P<0.01, compared with V23101 cells
图6 外源性TGFBI对裸鼠体内肿瘤组织中Ki67表达的影响
Figure 6 Effects of TGFBI on Ki67 expression in nude
mice tumor tissue in vivo |
TGFBI是TGF-β诱导分泌的一种细胞外分泌蛋 白(ECM),也被称为βig-H3。TGFBI参与多种 生理过程,包括细胞形态、黏附、转移、血管发 生及炎症[9]。小鼠胚胎组织的研究发现[10],胚胎 发育各阶段的许多组织间质都可以检测到TGFBI 的高表达,提示TGFBI对正常组织的发育是不可 缺少的。本研究也发现在永生化的乳腺上皮细胞 MCF-10F中,TGFBI蛋白不仅存在于培养细胞的 上清液中,也存在于细胞质和细胞核中,并且证 实了外源性TGFBI可以在乳腺癌MDA-MB-231细 胞中稳定性表达。Zhao等[4]发现在放射诱导的恶变 BEP2D细胞中TGFBI表达降低,过表达TGFBI后, 肿瘤细胞克隆形成率下降,同时裸鼠成瘤率也下 降,对130例人肺癌组织进行免疫组织化学分析发 现,有45个肺癌组织表现为TGFBI低表达。本研 究体外实验结果表明,外源性TGFBI可能抑制乳 腺癌细胞的增殖,并且显著地抑制了乳腺癌细胞 的锚着独立性生长能力;体内实验结果也发现, 外源性TGFBI可抑制乳腺癌细胞增殖,从而降低 乳腺癌发生率,这与以上研究结果相一致,提示 TGFBI对乳腺癌细胞的生长也存在抑制作用。但 是,TGFBI抑制肿瘤细胞生长的机制尚不明确。 Wen等[11]研究发现,TGFBI可以抑制肺癌和乳腺癌 细胞的体内外增殖,并且发现这种抑制作用与减 少软琼脂克隆形成能力、激活MMP2/9酶的活性, 从而提高细胞外基质穿透能力有关。
本研究结果显示,外源TGFBI能够使乳腺癌细 胞阻滞于G1期,从而减少进入S期的细胞,抑制肿 瘤细胞的增殖。有研究者发现[12],TGFBI缺陷小鼠 胚胎成纤维细胞更易发生染色体畸变,细胞增殖 活性增强,细胞复制过早进入S期,从而增加癌变 几率。这与本研究结论相符。TGFBI是否通过这 一途径调节细胞周期,还需要进一步研究。
细胞周期在G1期停留是为了检查DNA完整 性,主要是P53发挥作用。正常状态下,P53蛋白 含量低,当细胞异常时,p53基因活化,作用于 p21、DNA修复基因及Bax基因。p21基因活化后通 过抑制CDK及参与DNA复制的蛋白质,阻止细胞 进入S期,从而控制细胞增殖。本研究发现外源性 TGFBI可以使乳腺癌细胞P53和P21的瞬时表达增 高,与细胞周期停滞在G1期相一致,说明调控p53 和p21基因是TGFBI影响细胞周期的一个途径。但 精确的分子途径仍需进一步研究。
综上所述,TGFBI可以明显抑制乳腺癌细胞增 殖,作为乳腺癌治疗的肿瘤抑制基因,具有潜在 的应用价值。
参考文献
| [1] | Chen WQ,Zeng HM,Zheng RS,et al.Cancer incidence and mortality in China,2007[J].Chin J Cancer Res,2012,24 (1):1-8. |
| [2] | Siegel R,Naishadham D,Jemal A.Cancer statistics 2012[J].CA Cancer J Clin,2012,62(1):10-29. |
| [3] | Nam JO,Jeong HW,Lee BH,et al.Regulation of tumor angiogenesis by fastatin,the fourth FAS1 domain of betaig-h3,via alphavbeta3 integrin[J].Cancer Res,2005,65 (10):4153-61. |
| [4] | Zhao YL,Piao CQ,Hei TK.Overexpression of Betaig-h3 gene downregulates integrin alpha5beta1 and suppresses tumorigenicity in radiation-induced tumorigenic human bronchial epithelial cells[J]. Br J Cancer,2002,86 (12): 1923-8. |
| [5] | Zhao YL,Piao CQ,Hei TK.Downregulation of Betaig-h3 gene is causally linked to tumorigenic phenotype in asbestos treated immortalized human bronchial epithelial cells[J]. Oncogene,2002,21 (49):7471-7. |
| [6] | Zhao Y,El-Gabry M,Hei TK.Loss of Betaig-h3 protein is frequent in primary lung carcinoma and related to tumorigenic phenotype in lung cancer cells[J].Mol Carcinog,2006,45(2):84-92. |
| [7] | Ahmed AA,Mills AD,Ibrahim AE,et al.The extracellular matrix protein TGFBI induces microtubule stabilization and sensitizes ovarian cancers to paclitaxel[J].Cancer Cell,2007,12(6):514-27. |
| [8] | CaoYM,Zhang HM,Gao SH,et al.Transforming growth factor-β induced gene inhibits cell proliferation of malignant mesothelioma in vitro[J].Zhong Liu Fang Zhi Yan Jiu,2013,40(12):1123-7.[曹艳 梅,张鹤美,高四海,等.转化生长因子β诱导的基因抑制恶性间皮 瘤细胞增殖的体外研究[J].肿瘤防治研究,2013,40(12):1123-7.] |
| [9] | Thapa N,Lee BH,Kim IS.TGFBIp/betaig- h3 protein: a versatile matrix molecule induced by TGF-beta[J].Int J Biochem Cell Biol,2007,39 (12), 2183-94. |
| [10] | Schorderet DF,Menasche M,Morand S,et al.Genomic characterization and embryonic expression of the mouse Bigh3 (Tgfbi) gene[J]. Biochem Biophys Res Commun,2000,274(2),267-74. |
| [11] | Wen G,Partridge MA,Li B,et al.TGFBI expression reduces in vitro and in vivo metastatic potential of lung and breast tumor cells[J]. Cancer Lett,2011,308(1): 23-32. |
| [12] | Zhang Y,Wen G,Shao G,et al. TGFBI deficiency predisposes mice to spontaneous tumor development[J]. Cancer Res, 2009, 69(1): 37-44. |