 2014, Vol.41
2014, Vol.41



长链非编码RNA在肿瘤中的研究进展
引用本文
张彬彬,殷 媛,黄朝晖. 长链非编码RNA在肿瘤中的研究进展[J]. 肿瘤防治研究, 2014, 41(06): 669-673. 
ZHANG Binbin, YIN Yuan, HUANG Zhaohui. New Research of Long Noncoding RNA in Tumor. Cancer Research on Prevention and Treatment, 2014, 41(06): 669-673.
长链非编码RNA在肿瘤中的研究进展
214062 江苏无锡,江南大学附属医院 无
锡市肿瘤研究所
摘要:长链非编码RNA(Long noncoding RNAs,lncRNA)通常是指一类相对分子质量大于200 nt,
且不能翻译为蛋白质的调控性RNA分子。lncRNA在机体生理及病理过程中均具有广泛的功能,尤其
与恶性肿瘤发生发展关系密切。近年来lncRNA的研究进展迅速,本文就lncRNA在肿瘤中的最新研究
进展作一综述。
关键词:
长链非编码RNA
肿瘤
基因调控
New Research of Long Noncoding RNA in Tumor
Cancer Institute,The Affi liated Hospital of Jiangnan University,Wuxi 214062,China
Abstract:Long noncoding RNAs (LncRNAs) are regulatory non-coding RNA molecules which are longer than
200 nucleotides. LncRNA has a wide range of functions under physiological and pathological conditions, and
especially closely associated with the development of malignant tumor. LncRNA research has got signifi cant
advances in recent years. In this review, we summarize the recent research advances of LncRNA in cancer.
Key words:
LncRNA
Tumor
Gene regulation
0 引言
1.3 lncRNA的分子机制
哺乳动物基因组中蛋白质编码基因(protein coding gene,PCG)的比例不足2%,但剩下的巨 大的非蛋白编码区并非“垃圾DNA”,而是包含大 量转录调控元件和非编码RNA (noncoding RNA, ncRNA)基因。ncRNA通常是指不能翻译为蛋白 的RNA分子,包含短链ncRNA(如microRNA、 piRNA、siRNA等)和长链非ncRNA(lncRNA)。 目前发现或预测的人ncRNA已经超过了PCG,达到 4万以上,其中主要为lncRNA。lncRNA可在各个层 面调控基因表达,在细胞生长、发育、衰老、死亡 等过程中均扮演关键角色。 1 lncRNA概述 1.1 lncRNA的定义
lncRNA通常指一类相对分子质量大于200 nt, 且不能翻译为蛋白质的RNA分子。然而随着 研究的深入,发现该定义已经不能涵盖全部的 lncRNA。有学者建议将lncRNA定义为与传统类型 小RNA及结构RNA不同的,在初始转录或剪接后 水平调控基因的功能性RNA分子[ 1, 2 ]。 1.2 lncRNA的分类
目前lncRNA的分类和命名比较混乱,缺乏统 一标准[ 3 ]。可根据来源将其分为:(1)由PCG的基 因结构中断形成新的lncRNA;(2)染色质重排产 生出一个新的含有多个外显子的lncRNA;(3)非 编码基因在复制过程中由于反转座作用导致基因重 叠而产生的lncRNA;(4)局部的复制子串联产生 的lncRNA;(5)基因中插入一个转座元件而产生 的lncRNA。可根据在基因组定位的不同将lncRNA 分为:(1)反义lncRNA;(2)内含子lncRNA; (3)基因间lncRNA;(4)启动子相关lncRNA; (5)非翻译区lncRNA等。目前已经有很多在线数 据库开始收录lncRNA的相关信息,见表 1。
|
|
表 1 lncRNA公共数据库 Table 1 lncRNA in public databases |
lncRNA参与了基因组印迹、X染色体失活、 染色体修饰以及端粒生物学等多种生命活动过 程,参与人类多种疾病的发生和发展,尤其与恶 性肿瘤的发生、发展关系密切。由于lncRNA种类 较多,功能及作用机制复杂,目前尚知之甚少, 但也发现lncRNA的功能存在一定的共同点,可以 在转录水平、转录后水平以及表观遗传水平(如 染色质重塑、组蛋白修饰)调控基因表达。根据 现有的研究结果,研究者提出了一些作用模型, 鉴于已有中文综述提及[ 15 ],本文不再展开。 2 肿瘤相关lncRNA
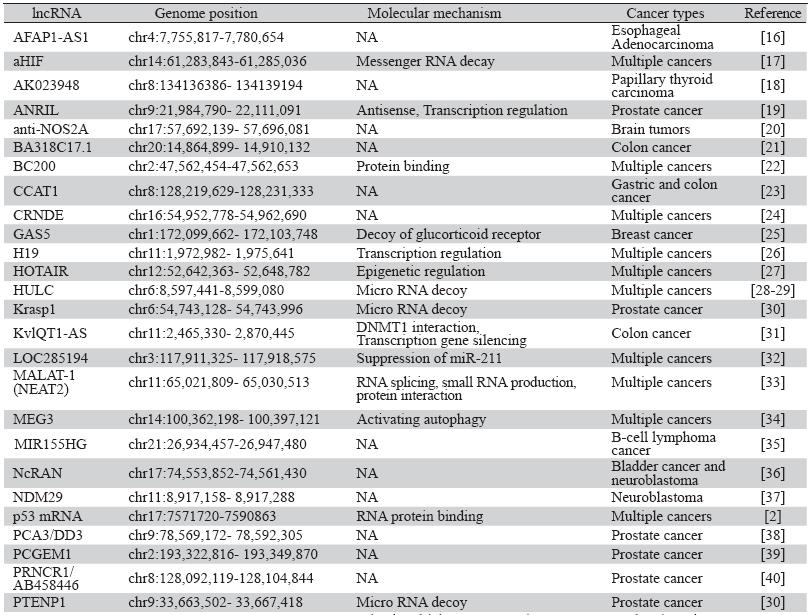
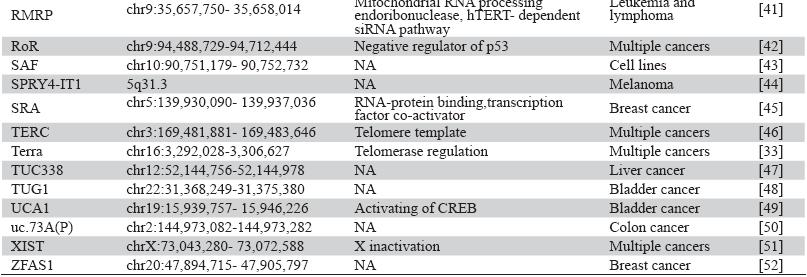
关于lncRNA的差异表达与肿瘤发生发展的 关系已经研究了很多年,然而,一直缺乏强有力 的直接证据。近年来随着肿瘤转录组表达谱的完 善,越来越多的证据揭示了lncRNA的功能,并发 现了大量的肿瘤相关lncRNA分子,见表 2。下面就 一些具有显著特点的肿瘤相关lncRNAs作一综述。
2.1 MALAT1这一大小约7.5 kb的lncRNA在人类多种肿瘤, 如肺癌、乳腺癌、胰腺癌、结肠癌以及前列腺癌 中异常高表达,其中尤以在非小细胞肺癌中的过 表达最为显著。研究已证实MALAT1的高水平表 达可促进肿瘤细胞转移。MALAT1的可能分子机 制包括剪接调节和基因表达调节。MALAT1可激 活转移相关基因的表达而促进肿瘤转移[ 33 ],还可 通过调节丝氨酸/精氨酸剪接因子的磷酸化水平使 mRNA前体的剪接发生改变[ 53 ],进而影响相应基因 的表达与功能。 2.2 HULC
HULC在肝癌和大肠癌肝转移中的表达显著增 高。HULC与乙肝病毒X蛋白HBx介导的肝癌发生 密切相关。HBx能够通过cAMP应答元件结合蛋白 (CREB)使HULC启动子激活[ 28 ]。此外,HULC 还可作为“分子诱饵”或是“miRNA海绵”在转录水 平下调miR-372的表达,导致其靶基因PRKACB水 平上调,激活CREB,CREB活化后与HULC启动 子结合从而促进HULC的转录[ 29 ]。 2.3 XIST
X染色体失活是哺乳动物表观遗传学调节的一 种经典模型,XIST是其主要调节因子。在雌性动 物细胞中,XIST可通过招募EZH2、RING-1等因 子到一条X染色体上,导致其失活,从而弥补了雌 雄性之间X相关基因数量的不平衡。乳腺癌中,可 存在两条活性X染色体。其共同机制是非活性X染 色体发生缺失或活性X染色体发生复制导致XIST 异常表达[ 51 ]。 2.4 HOTAIR
HOTAIR是第一个被证明可调控肿瘤转移的 lncRNA,在乳腺癌、肝细胞癌、喉鳞状细胞癌等 恶性肿瘤中表达显著上调,并与肿瘤的分期、转 移以及生存密切相关[ 27 ]。HOTAIR可募集PRC2复 合体并将其定位到HOXD位点,导致该位点上一大 小为40 kb的区域发生表达沉默,从而改变乳腺上 皮细胞的表型,使之类似于胚胎的纤维原细胞[ 54 ]。 HOTAIR还可能通过促进PTEN的甲基化导致其失 活进而促进肿瘤发生发展[ 55 ]。 2.5 GAS5
GAS5位于1q25,该区域在多种肿瘤中存在结 构异常。GAS5在缺乏营养或生长因子条件下大量 表达,扮演RNA Decoy角色,从而竞争抑制糖皮 质激素等类固醇激素介导的转录。GAS5在控制哺 乳动物细胞凋亡和细胞增殖过程中起重要作用, 可使乳腺癌细胞对凋亡诱导物更加敏感[ 25 ]。 2.6 H19
H19在脊椎动物胚胎发育过程中高表达,但出 生后不久即显著下调(除骨和软骨组织外)。H19 在多种肿瘤组织中异常高表达,可被癌基因c-Myc 直接活化,而抑癌基因p53则可下调其表达[ 26 ]。肝 脏中的H19与其作用靶点—血管生成素和成纤维细 胞生长因子之间相互作用,可改变与血管转移、 肿瘤浸润和细胞迁移相关基因的表达从而诱发肿 瘤。但也有证据表明H19可作为抑癌基因,如缺失 H19的小鼠较野生型更易发生肿瘤样息肉[ 56 ]。H19 同时兼有原癌基因和抑癌基因的作用,可能与其 特性相关,也可能与肿瘤的微环境有关[ 3 ]。 2.7 MEG3
这是第一个被发现具有肿瘤抑制功能的 lncRNA,其大小约1.6 kb。MEG3在人类很多正常 组织中表达,却在多种肿瘤组织中表达下调或丧 失,其机制包括基因缺失、启动子超甲基化等。 MEG3可以激活p53依赖或p53非依赖的信号通路抑 制肿瘤生成。由MEG3介导的p53的活化是依赖其 二级结构而不是其原始序列发挥作用的[ 34 ]。 2.8 UCA1
UCA1属于人内生性逆转录酶病毒H家族,在 绒膜绒毛、胚胎和胎儿膀胱组织中高表达。UCA1 在膀胱癌中表达上调,可促进膀胱癌细胞的增殖 和转移能力,并可通过CREB经由PI3K-AKT通路 调节细胞周期[ 49 ]。UCA1还可通过调控多个细胞周 期相关基因来改变细胞周期进程。 2.9 DD3
DD3基因位于9q21.22位点,目前尚未发现与 其同源的任何基因。现有资料表明DD3仅在前列 腺中特异性高表达,对前列腺癌的诊断价值要高 于目前常用的端粒酶逆转录酶,具有广阔的应用 前景[ 38 ]。 2.10 PTENP1
PTENP1是抑癌基因PTEN的假基因,与 其3 ’UTR高度相似,可竞争结合靶向PTEN的 miRNA(如miR-17、miR-21和miR-19等),从而解除 这些miRNA对PTEN的翻译抑制。PTENP1的发现 表明不同类型的ncRNA(miRNA和lncRNA)可共 同发挥基因调控功能[ 30 ]。 3 lncRNA在肿瘤诊断与治疗中的潜在应用
近年的研究显示,肿瘤的发生、发展、转移 与预后都与lncRNA相关。因此,lncRNA可作为 新的标志物用于肿瘤的诊断、预后和疗效监测。 例如,DD3用于前列腺癌诊断的敏感度和准确率 类似于PSA,且联合检测DD3和PSA可提高前列腺 癌的早期诊断率;检测尿沉渣UCA1用于膀胱癌的 诊断具有较高的特异性和敏感度;分析非小细胞 肺癌组织中MALAT1的含量可以早期预测肿瘤转 移;而HOTAIR是喉鳞状细胞癌、鼻咽癌的独立预 后因子[ 55 ]。
目前,将lncRNA用于肿瘤治疗干预靶点的研 究还处于起步阶段。尽管目前对lncRNA功能机制 所知有限,但已经为当前肿瘤药物的研发策略带来 很多提示,如合成可形成特定二级结构的RNA分 子,模拟lncRNA对基因的调控功能,抑制肿瘤的 增殖和转移。已经有部分lncRNA显示出作为肿瘤 干预靶点的巨大应用前景。例如,阻止HOTAIR与 PCR2复合物的反应,可限制乳腺癌细胞的转移潜 能。抑制GAS5表达不仅可诱导乳腺癌细胞的生长 停滞和凋亡,还可增加肿瘤对化疗药物的敏感度, 提示GAS5未来可能用于乳腺癌的预防和治疗[ 25 ]。 目前已经有一些靶向lncRNA的小分子化合物进入 了临床试验阶段。 4 展望
虽然lncRNA的研究已经进行了很多年,但 一直进展缓慢,总体上仍处于起步阶段,绝大多 数lncRNA的功能机制还不清楚。由于lncRNA种 类众多,结构多样,导致其作用机制复杂,目前 尚未建立成熟的研究技术体系,仅能对少数几类 lncRNA进行研究。此外,目前为止仍没有一个完 善的lncRNA数据库,仅建立了少数几个lncRNA转 基因模型。由于lncRNA广泛的生理功能和病理意 义,对lncRNA进行深入研究不仅会大大促进后基 因组时代对基因调控的理解,也必将对生物医学 的发展带来革命性的发展。
参考文献
| [1] | Spizzo R, Almeida MI, Colombatti A, et al. Long non-coding RNAs and cancer: a new frontier of translational research?[J].Oncogene,2012,31(43):4577-87. |
| [2] | Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions[J]. Nat Rev Genet,2009,10(3):155-9. |
| [3] | Gibb EA, Brown CJ, Lam WL. The functional role of long noncoding RNA in human carcinomas[J]. Mol Cancer,2011,10:38. |
| [4] | Erdmann VA, Szymanski M, Hochberg A, et al. Non-coding, mRNAlike RNAs database Y2K[J]. Nucleic Acids Res,2000,28(1):197-200. |
| [5] | Liu C, Bai B, Skogerb? G, et al. NONCODE: an integrated knowledge database of non-coding RNAs[J]. Nucleic Acids Res,2005,33(Database issue):D112-5. |
| [6] | Szymanski M, Erdmann VA, Barciszewski J. Noncoding RNAs database (ncRNAdb)[J]. Nucleic Acids Res,2007,35(Database issue):D162-4. |
| [7] | Mituyama T, Yamada K, Hattori E, et al. The Functional RNA Database 3.0: databases to support mining and annotation of functional RNAs[J]. Nucleic Acids Res,2009,37(Database issue):D89-92. |
| [8] | Zhang Y, Guan DG, Yang JH, et al. ncRNAimprint: a comprehensive database of mammalian imprinted noncoding RNAs[J]. RNA,2010,16(10):1889-901. |
| [9] | Dinger ME, Pang KC, Mercer TR, et al. NRED: a database of long noncoding RNA expression[J]. Nucleic Acids Res,2009,37(Database issue):D122-6. |
| [10] | Amaral PP, Clark MB, Gascoigne DK, et al. lncRNAdb: a reference database for long noncoding RNAs[J]. Nucleic Acids Res,2011,39(Database issue):D146-51. |
| [11] | Burge SW, Daub J, Eberhardt R,et al. Rfam 11.0: 10 years of RNA families[J]. Nucleic Acids Res 2013, 41(Database issue):D226-32. |
| [12] | Bejerano G, Pheasant M, Makunin I,et al. Ultraconserved elements in the human genome[J]. Science,2004,304(5675):1321-5. |
| [13] | Volders PJ, Helsens K, Wang X, et al. LNCipedia: a database for annotated human lncRNA transcript sequences and structures[J]. Nucleic Acids Res,2013,41(Database issue):D246-51. |
| [14] | Wu T, Wang J, Liu C,et al. NPInter: the noncoding RNAs and protein related biomacromolecules interaction database[J]. Nucleic Acids Res,2006,34(Database issue):D150-2. |
| [15] | Gao Y, Hui N, Liu S. Long Non-coding RNA:research progress[J]. Di Er Jun Yi Da Xue Xue Bao,2011,32(7):790-4.[高原, 惠宁, 刘善荣. 长链非编码RNA的研究进展[J]. 第二军医大学学报,2011,32(7):790-4.] |
| [16] | Wu W, Bhagat TD, Yang X, et al. Hypomethylation of noncoding DNA regions and overexpression of the long noncoding RNA, AFAP1-AS1, in barrett’s esophagus and esophageal adenocarcinoma[J]. Gastroenterology,2013,144(5):956-66.e4. |
| [17] | Rossignol F, Vaché C, Clottes E. Natural antisense transcripts of hypoxia-inducible factor 1alpha are detected in different normal and tumour human tissues[J]. Gene,2002,299(1-2):135-40. |
| [18] | He H, Nagy R, Liyanarachchi S, et al. A susceptibility locus for papillary thyroid carcinoma on chromosome 8q24[J]. Cancer Res,2009,69(2):625-31. |
| [19] | Kotake Y, Nakagawa T, Kitagawa K, et al. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of p15(INK4B) tumor suppressor gene[J]. Oncogene,2011,30(16): 1956-62. |
| [20] | Korneev SA, Korneeva EI, Lagarkova MA, et al. Novel noncoding antisense RNA transcribed from human anti-NOS2A locus is differentially regulated during neuronal differentiation of embryonic stem cells[J]. RNA,2008,14(10):2030-7. |
| [21] | Davison EJ, Tarpey PS, Fiegler H, et al. Deletion at chromosome band 20p12.1 in colorectal cancer revealed by high resolution array comparative genomic hybridization[J]. Genes Chromosomes Cancer,2005,44(4):384-91. |
| [22] | Chen W, B?cker W, Brosius J, et al. Expression of neural BC200 RNA in human tumours[J]. J Pathol,1997,183(3):345-51. |
| [23] | Yang F, Xue X, Bi J, et al. Long noncoding RNA CCAT1, which could be activated by c-Myc, promotes the progression of gastric carcinoma[J]. J Cancer Res Clin Oncol,2013,139(3):437-45. |
| [24] | Ellis BC, Molloy PL, Graham LD. CRNDE: A Long Non-Coding RNA Involved in CanceR, Neurobiology, and DEvelopment[J]. Front Genet,2012,3:270. |
| [25] | Mourtada-Maarabouni M, Pickard MR, Hedge VL, et al. GAS5, a non-protein-coding RNA, controls apoptosis and is downregulated in breast cancer[J]. Oncogene,2009,28(2):195-208. |
| [26] | Dugimont T, Montpellier C, Adriaenssens E, et al. The H19 TATA-less promoter is efficiently repressed by wild-type tumor suppressor gene product p53[J]. Oncogene,1998,16(18):2395-401. |
| [27] | Gupta RA, Shah N, Wang KC,et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis[J]. Nature,2010,464(7291):1071-6. |
| [28] | Du Y, Kong G, You X, et al. Elevation of highly up-regulated in liver cancer (HULC) by hepatitis B virus X protein promotes hepatoma cell proliferation via down-regulating p18[J]. J Biol Chem,2012,287(31):26302-11. |
| [29] | Wang J, Liu X, Wu H, et al. CREB up-regulates long non-coding RNA, HULC expression through interaction with microRNA-372 in liver cancer[J]. Nucleic Acids Res,2010,38(16):5366-83. |
| [30] | Poliseno L, Salmena L, Zhang J, et al. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology[J]. Nature,2010,465(7301):1033-8. |
| [31] | Tanaka K, Shiota G, Meguro M, et al. Loss of imprinting of long QT intronic transcript 1 in colorectal cancer[J]. Oncology,2001,60(3):268-73. |
| [32] | Liu Q,Huang J,Zhou N,et al.LncRNA loc285194 is a p53-regulated tumor suppressor[J]. Nucleic Acids Res,2013,41(9):4976-87. |
| [33] | Gutschner T, H?mmerle M, Eissmann M, et al. The non-coding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells[J]. Cancer Res,2013,73(3):1180-9. |
| [34] | Zhang X, Rice K, Wang Y, et al. Maternally expressed gene 3 (MEG3) noncoding ribonucleic acid: isoform structure, expression, and functions[J]. Endocrinology,2010,151(3):939-47. |
| [35] | Eis PS, Tam W, Sun L,et al. Accumulation of miR-155 and BIC RNA in human B cell lymphomas[J]. Proc Natl Acad Sci U S A,2005,102(10):3627-32. |
| [36] | Zhu Y, Yu M, Li Z,et al. ncRAN, a newly identified long noncoding RNA, enhances human bladder tumor growth, invasion, and survival[J].Urology,2011,77(2):510.e1-5. |
| [37] | Castelnuovo M, Massone S, Tasso R, et al. An Alu-like RNA promotes cell differentiation and reduces malignancy of human neuroblastoma cells[J]. FASEB J,2010,24(10):4033-46. |
| [38] | de Kok JB, Verhaegh GW, Roelofs RW, et al. DD3(PCA3), a very sensitive and specific marker to detect prostate tumors[J]. Cancer Res,2002,62(9):2695-8. |
| [39] | Ifere GO, Ananaba GA. Prostate cancer gene expression marker 1 (PCGEM1): a patented prostate- specifi c non-coding gene and regulator of prostate cancer progression[J]. Recent Pat DNA Gene Seq,2009,3(3):151-63. |
| [40] | Chung S,Nakagawa H,Uemura M,et al. Association of a novel long non-coding RNA in 8q24 with prostate cancer susceptibility[J]. Cancer Sci,2011,102(1):245-52. |
| [41] | Maida Y,Yasukawa M,Furuuchi M,et al. An RNA-dependent RNA polymerase formed by TERT and the RMRP RNA[J].Nature, 2009,461(7261):230-5. |
| [42] | Zhang A, Zhou N, Huang J, et al. The human long non-coding RNA-RoR is a p53 repressor in response to DNA damage[J]. Cell Res,2013,23(3):340-50. |
| [43] | Yan MD,Hong CC,Lai GM,et al. Identifi cation and characterization of a novel gene Saf transcribed from the opposite strand of Fas[J]. Hum Mol Genet,2005,14(11):1465-74. |
| [44] | Khaitan D, Dinger ME, Mazar J, et al. The melanoma-upregulated long noncoding RNA SPRY4-IT1 modulates apoptosis and invasion[J]. Cancer Res,2011,71(11):3852-62. |
| [45] | Lanz RB, McKenna NJ, Onate SA, et al. A steroid receptor coactivator, SRA, functions as an RNA and is present in an SRC-1 complex[J]. Cell,1999,97(1):17-27. |
| [46] | Cao Y, Bryan TM, Reddel RR. Increased copy number of the TERT and TERC telomerase subunit genes in cancer cells[J]. Cancer Sci,2008,99(6):1092-9. |
| [47] | Braconi C, Valeri N, Kogure T, et al. Expression and functional role of a transcribed noncoding RNA with an ultraconserved element in hepatocellular carcinoma[J]. Proc Natl Acad Sci U S A,2011,108(2):786-91. |
| [48] | Han Y, Liu Y, Gui Y, et al. Long intergenic non-coding RNA TUG1 is overexpressed in urothelial carcinoma of the bladder[J]. J Surg Oncol,2013,107(5):555-9. |
| [49] | Yang C,Li X,Wang Y,et al. Long non-coding RNA UCA1 regulated cell cycle distribution via CREB through PI3-K dependent pathway in bladder carcinoma cells[J]. Gene,2012,496(1):8-16. |
| [50] | Calin GA, Liu CG, Ferracin M, et al. Ultraconserved regions encoding ncRNAs are altered in human leukemias and carcinomas[J]. Cancer Cell,2007,12(3):215-29. |
| [51] | Sirchia SM, Tabano S, Monti L, et al. Misbehaviour of XIST RNA in breast cancer cells[J]. PLoS One,2009,4(5):e5559. |
| [52] | Askarian-Amiri ME, Crawford J, French JD, et al. SNORDhost RNA Zfas1 is a regulator of mammary development and a potential marker for breast cancer[J]. RNA,2011,17(5):878-91. |
| [53] | Tripathi V, Ellis JD, Shen Z, et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation[J]. Mol Cell,2010,39(6):925-38. |
| [54] | Tsai MC, Manor O, Wan Y,et al. Long noncoding RNA as modular scaffold of histone modifi cation complexes[J]. Science,2010,329 (5992):689-93. |
| [55] | Li D, Feng J, Wu T, et al. Long intergenic noncoding RNA HOTAIR is overexpressed and regulates PTEN methylation in laryngeal squamous cell carcinoma[J]. Am J Pathol,2013,182(1):64-70. |
| [56] | Yoshimizu T, Miroglio A, Ripoche MA, et al. The H19 locus acts in vivo as a tumor suppressor[J]. Proc Natl Acad Sci U S A,2008,105(34):12417-22. |