 2014, Vol.41
2014, Vol.41
氯化锂对白血病THP-1细胞生长的抑制作用及其对Wnt通路的影响
引用本文
许思苗,方 勇,周 密,赵雪娇,秦 宇,张沛凌,原瑞凤,
周剑峰,杨 漾. 氯化锂对白血病THP-1细胞生长的抑制作用及其对Wnt通路的影响[J]. 肿瘤防治研究, 2014, 41(06): 568-572. 
XU Simiao, FANG Yong, ZHOU Mi, ZHAO Xuejiao, QIN Yu,ZHANG Peiling,
YUAN Ruifeng, ZHOU Jianfeng, YANG Yang. Lithium Chloride Inhibits Proliferation of THP-1 Leukemia Cells and Activates Wnt
Signaling Pathway. Cancer Research on Prevention and Treatment, 2014, 41(06): 568-572.
氯化锂对白血病THP-1细胞生长的抑制作用及其对Wnt通路的影响
1. 430030 武汉,华中科技大学同济医学院
附属同济医院血液科
2.妇产科
2.妇产科
摘要:目的 研究氯化锂(LiCl)在体外对白血病细胞THP-1增殖、凋亡、周期以及Wnt通路的影
响。 方法 氯化锂不同浓度不同时间作用THP-1细胞后,用MTT法检测细胞增殖,流式细胞术检测
细胞凋亡和周期分布,Western blot检测细胞内Wnt通路的变化。 结果 氯化锂能抑制白血病THP-1细
胞的增殖,诱导细胞凋亡,使细胞周期阻滞于G2/M期,且均呈浓度和时间依赖性。氯化锂不同浓度
不同时间作用THP-1细胞后,t-GSK3β、cyclinD1表达不变,p-GSK3β、β-catenin表达升高,短时间低
浓度作用后c-myc表达升高,但随作用时间延长和浓度增加,c-myc表达降低。 结论 氯化锂对THP-1
细胞有显著的增殖抑制、凋亡诱导、细胞G2/M期阻滞作用,并激活Wnt通路,影响Wnt通路下游蛋白
c-myc、cyclinD1的表达。
关键词:
氯化锂
白血病
THP-1细胞系
增殖
凋亡
细胞周期
糖原合成酶激酶
Wnt信号通路
Lithium Chloride Inhibits Proliferation of THP-1 Leukemia Cells and Activates Wnt Signaling Pathway
XU Simiao1, FANG Yong2, ZHOU Mi1, ZHAO Xuejiao2, QIN Yu2,ZHANG Peiling1, YUAN Ruifeng1, ZHOU Jianfeng1, YANG Yang1

1.Department of Hematology,Tongji Hospital,Tongji Medical College,Huazhong University of
Science and Technology,Wuhan 430030,China
2. Department of Gynaecology and Obstetrics
2. Department of Gynaecology and Obstetrics
Abstract:Objective To explore the effects of Lithium Chloride (LiCl) on proliferation, apoptosis and cell
cycle of leukemia cells THP-1, and to investigate its effect on Wnt signaling pathway. Methods THP-1 cells
were treated with LiCl at different concentrations and time. The proliferation inhibitory rates of THP-1 cells
were assayed by MTT. The apoptosis rates and the cell cycle distribution were detected by fl ow cytometry.
The expressions of Wnt signaling pathway proteins were analyzed by Western blot. Results LiCl could
inhibit the proliferation and induce the apoptosis of THP-1 cells. LiCl mainly arrested THP-1 cells in the
G2/M phase in a time- and dose-dependent manner. Western blot results suggested that the expressions of
t-GSK3β and cyclinD1 had no obvious change. The expressions of p-GSK3β and β-catenin were increased.
The expression of c-myc protein was increased with short-time and low-concentration treatment, while
decreased with long-time and high-concentration treatment. Conclusion LiCl could inhibit proliferation,
induce apoptosis and arrest cell cycle of THP-1 cells. In addition, it could activate Wnt signaling pathway, and
infl uence the expressions of c-myc and cyclinD1 proteins.
Key words:
Lithium chloride
Leukemia
THP-1 cell line
Proliferation
Apoptosis
Cell cycle
GSK-3
Wnt signaling pathway
0 引言
2.2 氯化锂对THP-1细胞凋亡率的影响
2.3 氯化锂对THP-1细胞周期分布的影响
2.4 氯化锂对THP-1细胞Wnt通路的影响
3 讨论
氯化锂(lithium chloride,LiCl)是治疗躁郁 型精神障碍的主要药物,是糖原合成酶激酶-3 (GSK-3)的抑制剂。大量研究显示,氯化锂在人体多个系统中具有广泛的生理调节作用,在神经 变性、糖尿病、中风、肿瘤、慢性炎症等疾病中 有一定的治疗意义[ 1 ];在造血系统中能增强造血干 细胞的自我更新和增殖分化能力,临床上用于放 化疗后造血系统的恢复[ 2 ],相关机制研究发现,这 些作用均与其对GSK-3的抑制密切相关[ 3 ]。但氯化 锂对白血病细胞THP-1的作用,目前尚未见报道, 本文研究了氯化锂对THP-1细胞增殖、凋亡、周期 的影响,并初步观察了氯化锂作用后细胞内Wnt通 路的变化情况。 1 材料与方法 1.1 材料
人THP-1白血病细胞株购自美国ATCC公司。 RPMI1640培养液购自美国Gibco公司,胎牛血清 购自杭州四季青公司。氯化锂购自美国Sigma公 司。噻唑蓝(MTT)、二甲基亚砜(DMSO)、 碘化吡啶(PI)购于美国Sigma公司。Annexin V-FITC/PI双染试剂盒购自南京凯基生物公司。 GSK3β、P-GSK3β(Ser9)、β-catenin、c-myc、 cyclinD1兔抗人单克隆抗体均购自美国C e l l Signaling公司,GAPDH鼠抗人单克隆抗体购自美 国Epitomics公司。皮克级化学发光底物试剂盒购 自美国Thermo公司。 1.2 细胞增殖MTT实验
将细胞(1×104 /ml)接种于96孔板,200微升 /孔,加入终浓度为0、2、 4、 8、 16、 32、64及 128 mM/ml的氯化锂,以不加细胞和药物的培养 基为空白对照,每组设4个平行复孔。分别于培养 后24、48、72、96 h后,每孔加入5 mg/ml MTT20 μl,继续培养4 h,离心去除上清,每孔加入200 μl DMSO,振荡溶解,酶标仪492 nm波长处测吸光 度(A),按如下公式计算抑制率。
抑制率%=[1- ( A实验组 - A空白对照组 ) / (A阴性对照组 - A空白对照组 )]×100% 1.3 流式细胞术检测细胞凋亡
氯化锂各浓度作用THP-1细胞24、48 h后,收 集细胞,PBS洗涤两次后,取1×106细胞,用500 μl 的Binding Buffer重悬,加入2 μl Annexin V-FITC 和 5 μl PI,混匀,室温避光反应15 min后,于1 h内上 机检测。 1.4 流式细胞术检测细胞周期分布
收集氯化锂各浓度处理24、48 h细胞,PBS洗 涤两次,取5×105细胞离心后沉淀物,加入70%冰 乙醇-20℃过夜。离心去上清,沉淀用200 μlPBS 重悬,加入20 μlRNA酶,室温5 min,再加入50 μl PI,室温避光20 min,于1 h内上机检测。 1.5 Wnt通路的Western blot检测
收集氯化锂各浓度处理4、12、24、48 h的细 胞,用RIPA蛋白裂解液提取收集细胞蛋白质,用 考马斯亮蓝法测定蛋白浓度。取80 μg变性后蛋白 样品上样于10%SDS-PAGE,电泳后冰浴电转至 PVDF膜上,5%脱脂奶粉室温封闭1 h,加入各一 抗抗体4℃孵育过夜。TBS洗膜后,加入荧光二抗 室温孵育1 h。TBS洗膜,用皮克级化学发光底物 试剂盒进行曝光显影。 1.6 统计学方法
结果数据以均数±标准差(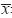 ±s)表示,应用
GraphPad Prism 5.0 软件进行统计分析,t检验分析
实验组和对照组之间差异,以P<0.05为差异具有
统计学意义。
2 结果
2.1 氯化锂对THP-1细胞增殖的影响
±s)表示,应用
GraphPad Prism 5.0 软件进行统计分析,t检验分析
实验组和对照组之间差异,以P<0.05为差异具有
统计学意义。
2 结果
2.1 氯化锂对THP-1细胞增殖的影响
MTT法检测显示,8~128 mM/ml氯化锂作 用THP-1细胞24、48、72、96 h,其抑制率均 明显高于对照组(8 mM/ml24、48、72 h时, P=0.0079、P=0.0079、P=0.0076;8 mM/ml作用 96 h以及16~128mM/ml 作用24、48、72、96 h,P 均<0.0001),且抑制率随浓度增大和时间延长而 增加。当4 mM/ml氯化锂作用白血病THP-1细胞96 h,其抑制率也明显高于对照组(P=0.0002,), 见图 1。
 |
图 1 氯化锂对THP-1细胞增殖的影响 Figure 1 Effects of LiCl on proliferation of THP-1 cells |
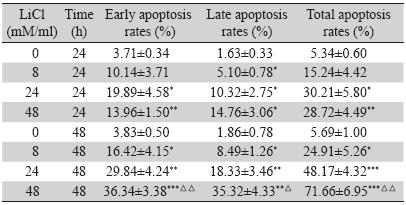
以流式散点图右下象限的早期凋亡细胞与 右上象限的晚期凋亡细胞之和表示总凋亡率。 结果显示,氯化锂8 mM/ml作用24 h,细胞凋 亡率与对照组相比,差异无明显统计学意义 (P=0.0908),而作用48 h后,细胞凋亡率明显高 于对照组(P=0.023);24和48 mM/ml氯化锂分别 作用24、48 h,细胞凋亡率明显高于对照组(24 mM/mL作用24、48 h ,P=0.013、P=0.0007,48 mM/ml作用24、48 h,P=0.0067、P=0.0007);且 48mM/ml氯化锂作用48 h与作用24 h相比,凋亡率 明显增高,差异有统计学意义(P=0.0066),见 表 1、图 2。
|
|
表 1 流式细胞术检测氯化锂作用THP-1细胞后的凋亡率(±s,
n=3)
Table 1 Effects of LiCl on apoptosis rates of THP-1 cells
detected by fl ow cytometry(±s,n=3) |
 |
A,B,C,D: 0,8,24,48mM/ml treatment for 24h,respectively; E,F,G,H: 0,8,24,48mM/ml treatment for 48h,respectively 图 2 THP-1细胞经氯化锂处理后的凋亡检测 Figure 2 Effects of LiCl on apoptosis of THP-1 cells |
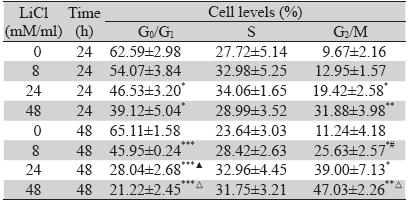
流式细胞术检测细胞周期分布,结果显示: 氯化锂8 mM/ml作用细胞24 h,细胞周期变化不 明显(G0/G1期P=0.1543,S期P=0.514,G2/M期 P=0.2887),8 mM/ml作用48 h、24和48 mM/ml 分别作用24和48 h,与对照组相比,G0/G1期细胞 比例明显减少(8 mM/ml作用48 h,P=0.0003, 24 mM/ml作用24、48 h,P=0.0214、P=0.0003; 48 mM/ml作用24、48 h,P=0.0161、P=0.0001), G2/M期细胞比例明显增多(8 mM/ml作用48 h, P=0.0427;24 mM/ml作用24、48 h,P=0.0446、 P=0.0283;48 mM/ml作用24、48 h,P=0.0081、 P=0.0017),S期细胞比例变化不明显(8 mM/ml 作用48 h,P=0.2997;24 mM/ml作用24、48 h, P=0.3058、P=0.1587;48 mM/ml作用24、48 h, P=0.8481、P=0.1404);48 mM/ml作用细胞48 h和 作用24 h相比,G0/G1、G2/M期细胞比例均差异显 著(P=0.0331,P=0.0297)。提示氯化锂将THP-1 细胞阻滞于G2/M期,且浓度越大、时间越长,阻 滞越明显,见表 2、图 3。
|
|
表 2 流式细胞术检测氯化锂作用THP-1细胞后的周期分布
(±s,n=3)
Table 2 Effects of LiCl on cell cycle of THP-1 cells detected
by fl ow cytometry(±s,n=3) |
 |
A,B,C,D: 0,8,24,48mM/ml treatment for 24h,respectively; E,F,G,H: 0,8,24,48mM/ml treatment for 48h,respectively 图 3 THP-1细胞经氯化锂处理后的周期检测 Figure 3 Effects of LiCl on cell cycle of THP-1 cells |
氯化锂8、24、48 mM/ml作用于THP-1细胞4、 12、24、48 h,用Western blot检测Wnt通路各蛋白 表达情况,结果显示,与对照组相比,t-GSK3β在 各浓度各时间作用下,表达基本不变;p-GSK3β表 达升高;β-catenin随浓度增加和时间延长而升高; 氯化锂各浓度作用4和12 h时,β-catenin下游c-myc 表达升高,8和24 mM/ml作用24h,以及8 mM/ml作 用48 h时,c-myc蛋白表达也升高,而在48mM/ml作 用24 h,24和48 mM/ml作用48 h时,c-myc蛋白表达 则下调;β-catenin下游另一蛋白cyclinD1则在氯化 锂各浓度各时间作用下表达变化不明显,见图 4。
 |
图 4 Western blot检测THP-1细胞经氯化锂作用后Wnt信号通路的变化 Figure 4 Effects of LiCl on expressions of Wnt signaling pathway proteins in THP-1 cells |
糖原合成酶激酶-3(GSK-3)是一种多功能 丝/苏氨酸蛋白激酶,普遍存在于所有真核细胞 生物,在多个信号转导通路中扮演重要角色,影 响细胞的基因表达、蛋白合成、糖原代谢以及增 殖、凋亡等生理过程,GSK-3的异常调节可能导 致糖尿病、心血管疾病、神经精神疾病等多种人 类疾病的发生[ 4 ]。以及肿瘤的发生与进展,其对肿 瘤调控最重要的一条途径是参与Wnt/β-catenin信号 通路。Wnt通路活化能抑制 GSK-3活性,使胞浆 内β-catenin水平升高并发生核转移,进入胞核的 β-catenin与转录因子TCF/LEFs结合,从而刺激下 游促癌基因(c-myc、cyclinD1、MMP-7)的高表 达,由此推测,抑制GSK-3可能促进肿瘤生长[ 5 ]。 但是,事实并非如此,有研究证实,在GSK3β缺 失的小鼠中,β-catenin和cyclinD1水平并不升高; 在结肠腺瘤性息肉病基因(APC)突变的小鼠中使用 GSK-3抑制剂氯化锂并不能导致肿瘤的发生;还有 研究表明长期使用氯化锂治疗精神疾病的人群肿 瘤发生率低于普通人群。因此,GSK-3对肿瘤是促 进还是抑制,目前仍存争议,一方面,GSK-3通 过对β-catenin的调节抑制肿瘤发生,但另一方面, GSK-3抑制剂却并不增加肿瘤发生率,相反对肿 瘤有一定的抑制作用,这可能与GSK-3参与除Wnt/ β-catenin通路以外的其他调节途径有关[ 6 ]。
多个研究表明,在血液系统恶性肿瘤,如慢 性粒细胞白血病(CML)、急性淋巴细胞白血病 (ALL)和急性髓系白血病(AML)中,均能检 测到Wnt通路的活化[ 7 ]。也有文献报道,GSK-3 抑制剂BIO对白血病细胞系有生长抑制作用[ 8, 9 ] , GSK-3是急性髓系白血病(AML)一个潜在的治 疗靶点[ 10 ]。然而,GSK-3抑制剂氯化锂对白血病 细胞系THP-1的生长以及细胞内Wnt通路的影响, 目前还未有相关研究。本实验中,氯化锂能抑制 THP-1细胞的增殖,且具有浓度和时间依赖性, 流式细胞术检测发现,不同浓度的氯化锂作用于 THP-1细胞24和48 h,细胞凋亡率均明显高于对照 组。此外,氯化锂对THP-1细胞的周期影响显著, 在不同浓度和时间的处理下,细胞阻滞于G2/M 期,S期变化不明显,G0/G1期细胞相对减少。对细 胞内Wnt通路进行Western blot检测,可以发现,不 同浓度氯化锂分别作用THP-1细胞4、12、24、48 h,Wnt通路活化,β-catenin随着作用时间和浓度 的增加表达明显升高,其下游c-myc蛋白在作用时 间短,浓度低时表达升高,但随着作用时间的延 长和浓度的增加,其表达也逐渐降低,而另一下 游分子cyclinD1则表达不变。
由此推测,在THP-1细胞中,氯化锂有效抑 制了GSK-3的活性,使得Wnt通路活化,β-catenin 升高,在短时间和低浓度情况下,促进下游靶基 因c-myc的表达,使得c-myc水平升高,c-myc是 细胞周期和凋亡相关蛋白,能促进细胞进入细胞 周期和发生凋亡[ 11,12 ] ,因此在本实验中,氯化锂短 时间低浓度的刺激即可促进THP-1细胞从G0/G1期 进入G2/M期,并同时发生凋亡。然而,随着作用 时间和浓度的增加,抑制GSK-3可能通过其他途 径抑制c-myc表达,从而使得c-myc下调,细胞发 生周期阻滞和凋亡[ 13,14 ] ,因此THP-1细胞被阻滞 于G2/M期,凋亡亦可同时发生。作为重要的肿瘤 调控蛋白[ 15 ],c-myc的表达变化可能是氯化锂抑 制白血病细胞THP-1生长的机制之一。另外,当 β-catenin升高,cyclinD1可能受其调控升高,但同 时,抑制GSK-3也能抑制NF-κB途径,使其下游 的cyclinD1下调,因此,氯化锂作用THP-1细胞可 能导致cyclinD1表达水平不变[ 5,6 ]。 当然,c-myc和 cyclinD1参与了细胞复杂的周期和凋亡调控系统, 氯化锂对THP-1细胞周期和凋亡的影响尚不能仅通 过c-myc和cyclinD1的变化来解释,还有待进一步 的研究。
总之,虽然GSK-3对肿瘤的调控机制尚不明 朗,但本实验证明了GSK-3抑制剂氯化锂能抑制 白血病细胞THP-1的生长,促进细胞凋亡和诱导 G2/M期阻滞,发挥抗白血病作用,为进一步探索 氯化锂在白血病中的作用机制,以及临床治疗白 血病的可行性提供了一定的研究基础。
参考文献
| [1] | Gould TD, Gray NA, HK. Effects of a glycogen synthase kinase-3 inhibitor, lithium, in adenomatous polyposis coli mutant mice[J].Pharmacol Res, 2003, 48(1): 49-53. |
| [2] | Huang J, Zhang Y, Bersenev A, et al. Pivotal role for glycogen synthase kinase-3 in hematopoietic stem cell homeostasis in mice[J]. J Clin Invest, 2009, 119(12): 3519-29. |
| [3] | Grimes CA, Jope RS. The multifaceted roles of glycogen synthase kinase 3beta in cellular signaling[J]. Prog Neurobiol, 2001, 65(4): 391-426. |
| [4] | Rayasam GV, Tulasi VK, Sodhi R, et al. Glycogen synthase kinase 3: more than a namesake[J]. Br J Pharmacol, 2009, 156(6): 885-98. |
| [5] | Luo J. Glycogen synthase kinase 3beta (GSK3beta) in tumorigenesis and cancer chemotherapy[J]. Cancer Lett, 2009, 273(2): 194-200. |
| [6] | Ougolkov AV, Billadeau DD. Targeting GSK-3: a promising approach for cancer therapy?[J]. Future Oncol, 2006, 2(1): 91-100. |
| [7] | Ge X, Wang X. Role of Wnt canonical pathway in hematological malignancies[J]. J Hematol Oncol, 2010, 3:33. |
| [8] | Song EY, Palladinetti P, Klamer G, et al. Glycogen synthase kinase--3β inhibitors suppress leukemia cell growth[J]. Exp Hematol, 2010, 38(10): 908-21. |
| [9] | Holmes T, O’Brien TA, Knight R, et al. Glycogen synthase kinase-3beta inhibition preserves hematopoietic stem cell activity and inhibits leukemic cell growth[J]. Stem Cells, 2008, 26(5): 1288-97. |
| [10] | Banerji V, Frumm SM, Ross KN, et al. The intersection of genetic and chemical genomic screens identifies GSK-3α as a target in human acute myeloid leukemia[J]. J Clin Invest, 2012, 122(3): 935-47. |
| [11] | Dang CV. c-Myc target genes involved in cell growth, apoptosis, and metabolism[J]. Mol Cell Biol, 1999, 19(1): 1-11. |
| [12] | Shortt J, Johnstone RW. Oncogenes in cell survival and cell death[J]. Cold Spring Harb Perspect Biol, 2012, 4(12). |
| [13] | Ryves WJ, Harwood AJ. The interaction of glycogen synthase kinase-3 (GSK-3) with the cell cycle[J]. Prog Cell Cycle Res, 2003, 5:489-95. |
| [14] | Cornils H, Kohler RS, Hergovich A, et al. Downstream of human NDR kinases: impacting on c-myc and p21 protein stability to control cell cycle progression[J]. Cell Cycle, 2011, 10(12): 1897-904. |
| [15] | Albihn A, Johnsen JI, Henriksson MA. MYC in oncogenesis and as a target for cancer therapies[J]. 2010, 107:163-224. |