 2014, Vol.41
2014, Vol.41


集落刺激因子-1及其受体对乳腺癌荷瘤鼠肿瘤生长的影响
引用本文
郝 娜,李盈盈,李 宁,张 为,阿卜杜凯尤木·麦麦提,杨玉姣,郑慧琳,王 悦. 集落刺激因子-1及其受体对乳腺癌荷瘤鼠肿瘤生长的影响[J]. 肿瘤防治研究, 2014, 41(06):536-540. 
HAO Na, LI Yingying, LI Ning,ZHANG Wei,Abudukayum·Maimaiti,YANG Yujiao, ZHENGHuilin, WANG Yue. Infl uence of Colony Stimulating Factor-1 and Its Receptor on Cancer Cells and Immune Cells of Breast Cancer Mice. Cancer Research on Prevention and Treatment, 2014, 41(06): 536-540.
集落刺激因子-1及其受体对乳腺癌荷瘤鼠肿瘤生长的影响
300071 天津, 南开大学医学院免疫学教研室
摘要:目的 探讨CSF-1及其受体对乳腺癌荷瘤鼠肿瘤体积以及对其免疫器官包括骨髓、胸腺、淋巴
结和脾的影响。方法 将4T1细胞(1×107/只)重悬于PBS液接种于Balb/c 小鼠左腋皮下,建立荷乳腺
癌小鼠模型为实验组,同时培养相同条件的对照组。检测成瘤率,测量肿瘤体积的变化并绘制成图;
成瘤后为实验组小鼠注射CSF-1及其受体,观察一段时间后同时处死两组小鼠,采用半定量RT-PCR技
术检测鼠脾脏中CSF-1的表达;采用MTT比色法检测小鼠的肿瘤、脾脏、骨髓中细胞的增殖功能。 结果 4T1细胞接种后小鼠的成瘤率为100%,成瘤潜伏期平均为7~10天;实验组小鼠瘤体的增长速度
快于对照组;RT-PCR的电泳结果显示,实验组小鼠的脾脏CSF-1的mRNA的表达量上升;MTT结果
表明,CSF-1作用之后的实验组小鼠的肿瘤细胞和免疫细胞增殖活性升高,发挥了强大的免疫抑制功
能,促进肿瘤的增长;增殖活跃的骨髓和脾细胞呈现免疫抑制表型。结论 CSF-1及其受体对乳腺癌
荷瘤鼠肿瘤的生长有促进作用。
关键词:
4T1乳腺癌细胞
集落刺激因子-1及其受体
肿瘤免疫
Infl uence of Colony Stimulating Factor-1 and Its Receptor on Cancer Cells and Immune Cells of Breast Cancer Mice.
Department of Immunology, Nankai University School of Medicine, Nankai University,Tianjin 300071, China
Abstract:Objective To study the influence of colony stimulating factor-1(CSF-1) and its receptor on
cancer cells and immune cells of breast cancer mice and to explore their effects on tumor size, volume and
immune system including marrow, thymus, lymphonodus and spleen. Methods4T1 cell (1×107/PBS) was
inoculated in the left subcutaneous of Balb/c mice in order to establish the mice model with breast cancer.
Then tumor formation rate, average weight and area change of tumor were measured. CSF-1 and its receptor
were injected into experimental mice after tumor was formed, and these mice were killed after observation.
The expression of CSF-1 in the spleen of mice was tested by semi-quantitative RT-PCR. The proliferation
of T lymphocyte in the spleen and bone marrow were detected by MTT colorimetry. ResultsAfter the
continuous improvement, tumor formation rate of 4T1 breast cancer was 100% and tumor latency was 7-10
days. After the injection of CSF-1, we found that tumor growth rate of the experimental mice was higher than
that of control group. RT-PCR and electrophoresis results revealed that expression of CSF-1 was increased
in the spleen of tumor-bearing mice.MTT results showed that the proliferation of immune lymphocytes
and tumor cells was increased after CSF-1 treatment. Active proliferation of bone marrow and spleen cells
exhibited immunosuppressive phenotype. Conclusion CSF-1 and its receptor could promote the growth of
breast cancer tumor in mice.
Key words:
4T1 breast cancer cell
Colony stimulating factor-1 and its receptor
Tumor immunity
0 引言
1.2.2 MTT比色实验
2.2 细胞检测
CSF-1及其受体作用后肿瘤细胞和免疫细胞
均表现增殖活性增加
2.2.2 CSF-1及其受体作用后荷瘤鼠肿瘤细胞中
CSF-1及相关癌基因的mRNA表达增高
2.2.3 CSF-1及其受体注射后荷瘤鼠免疫细胞中抑
制表型分子mRNA表达增高
3 讨论
乳腺癌的致癌机制可能是多步骤、多因素、 多基因协同或单独作用于机体不同阶段的复杂过 程,在正常情况下,机体存在多种原癌基因并参 与细胞的生长、增殖、分化以及信号转导,但在 异常情况下,当原癌基因非正常表达或过表达则 会导致相应的细胞增殖分化异常甚至成为克隆性生长的肿瘤细胞。而由原癌基因c-fms编码的集落 刺激因子CSF-1的受体CSF-1R的过度表达对乳腺 癌肿瘤的发生、发展起很重要的作用[1]。
CSF-1是体内重要的细胞因子,在进行造血 细胞体外研究中,它可刺激不同的造血干细胞在 半固体培养液中形成细胞,在体外实验中对骨髓 和脾巨噬细胞系有刺激作用[2]。而集落刺激因子 CSF-1受体是c-fms基因编码的产物,它是一种酪 氨酸激酶受体,当它与其配体CSF-1结合后会激活 受体酪氨酸激酶信号系统,并使酪氨酸激酶的活 性持续升高,从而改变效应器的生物活性,使相 应的细胞持续生长增殖,并且对细胞还发挥长时 效的作用,最后分化发生癌变[3, 4]。
本研究拟通过建立乳腺癌荷瘤鼠动物模型, 探讨CSF-1及其受体对乳腺癌荷瘤鼠肿瘤的作用 及对免疫器官脾和骨髓的影响。实验检测小鼠肿 瘤的体积大小变化,探其相关基因的表达,以及 检测鼠肿瘤、脾脏和骨髓细胞的增殖生长活性, 对鼠的胸腺、脾和淋巴结的细胞进行表型分析鉴 定,从而更加全面地研究探讨CSF-1及受体对乳腺 癌荷瘤鼠肿瘤的生长和机体免疫的作用。 1 材料与方法 1.1 材料
Balb/c小鼠,雌性,250~300 g,由中国人民 解放军军事医学研究院实验动物中心提供;4T1 细胞购自中国医学科学院肿瘤医院;Trizol,DNA marker购自北京普博欣生物科技公司 ;RPMI 1640 培养液购自美国Gibco公司;GAPDH,引物由美 国Invitrogen公司合成;一步法RT-PCR试剂盒购 自美国Bioer公司;MTT试剂购自天津润泰科技有 限公司;CSF-1(rM-CSF)购自美国Peprotech 公 司;CSF-1受体(CSF1R/MCSF Receptor)购自北京 义翘神州生物技术有限公司(Sino Biological Inc.)。 1.2 方法 1.2.1 RT-PCR
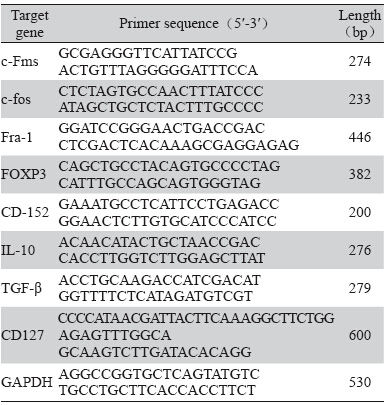
RT-PCR技术是目前用于对提取的目标RNA进 行反转录的方法。本实验设计了如下的引物分别 对小鼠脾细胞进行反转录实验,测定CSF-1作用后 细胞相关目的基因mRNA的表达,见表 1。 反应参数:50℃反转录30 min,94℃预热5 min后,94℃ 30 s,47℃ 30 s,72℃ 3 min,循环 40次,72℃延伸5 min。用Originpro8.5软件处理 数据,以引物因子为横坐标,以所测OD值为纵坐 标,获得相关函数,绘制标准曲线。
|
|
表 1 相关目标基因RT-PCR的引物序列 Table 1 RT-PCR associated primer sequences of target gene |
利用MTT比色法测定免疫器官包括脾细胞 和骨髓细胞,及其肿瘤细胞对刺激物CSF-1的反 应,进而来推断细胞增殖活性功能的变化。用 Originpro8.5软件来处理数据,绘制曲线。 1.2.3 动物实验
在无菌条件下,为对照组(A组)和实验组(B组) 左侧腋下注射1×107浓度的4T1细胞悬液建立荷瘤 鼠模型,于注射后第7天成瘤,并检测小鼠的成瘤 率。在瘤体出现的第4天为实验组小鼠注射CSF-1 及其受体,每天注射rM-CSF 3~4 ml (200 ng/ml); CSF1R/MCSF Receptor 50×104~100×104IU/次连 续2天,对照组小鼠注射相同剂量的0.9%氯化钠溶 液。从成瘤的第1天开始,记录肿瘤长径(L)和短 径(W),每天早中晚各一次取其平均值,用Steel 公式:V=LW2/2计算肿瘤的体积。于成瘤后第8天 断颈法处死小鼠取出肿瘤、骨髓和脾脏,做细胞实 验,将对照组与实验组的三种标本(脾,骨髓,肿 瘤)的新鲜细胞分于两份,第一份以Trizol法提取 总RNA,做RT-PCR和电泳,第二份培养24 h,做 MTT实验。目的在于测定CSF-1及其受体刺激后目 的mRNA的表达变化和各成分细胞活性增殖情况。 1.3 统计学方法
采用SPSS17.0软件进行统计分析,计量数据 以均数±标准差(x±s),多个样本均数多重比较采用 方差分析及SNK-q检验,以双侧检验P<0.05为差 异有统计学意义。 2 结果 2.1 动物实验结果
术后同期放化疗组生存率明显高于单纯放射 在未作用CSF-1及其受体的前三天内,实验组 的肿瘤体积增长速度为0.0357,对照组的增长速度 为0.0374,两者相比差异无统计学意义(P>0.05), 而CSF-1及其受体作用后实验组肿瘤增长速度为 0.0216,对照组的增长速度为0.0091,实验组是 对照组的2.4倍,两组相比差异有统计学意义(P <0.05),见图1。
 |
图 1CSF-1及其受体对肿瘤体积的影响 Figure 1Effects of CSF-1 and its receptor on tumor volume |
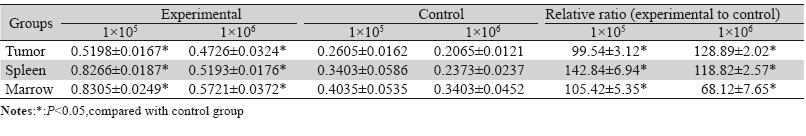
用MTT实验检测CSF-1及其受体作用后荷 瘤鼠肿瘤细胞、脾细胞及骨髓细胞的增殖生长 活性。实验首先在1×106的细胞浓度下进行细胞 培养,为了验证不同浓度的细胞数目是否会对 MTT的结果有影响,我们又用1×105的细胞进行 实验,检测各细胞活性。结果为在1×105浓度下, 实验组肿瘤细胞、脾细胞及骨髓细胞的活性均高 于对照组,由统计学分析得出其相对比分别为 (99.54±3.12)%、(142.84±6.94)%、(105.42 ±5.35)%(P<0.05),其相对值倍数分别为2.0、 2.4、2.1,结果均具有统计学意义。在1×106浓度 下,实验组上述三种细胞活性同样均高于对照 组,由统计学分析得其相对比分别为(128.89± 2.02)%、(118.82±2.57)%、(68.12±7.65)% (P<0.05),其相对值倍数为2.3、2.2、1.7,结果 具有明显的统计学意义。因此MTT实验结果表明 CSF-1及其受体作用后小鼠的肿瘤细胞增殖活跃, 同时脾细胞、骨髓细胞都有增殖活性升高的表 现,见表 2、图 2。
|
|
表 2 小鼠MTT实验细胞活性检测(x±s,%) Table 2 Proliferation of mice tumor and lymphocyte detected by MTT assay(x±s,%) |
 |
图 2MTT实验检测细胞增殖活性的结果 Figure 2Cell proliferation activity determined by MTT test |
为了研究CSF-1及其受体对荷瘤鼠肿瘤生长的 影响,我们收集了未治疗荷瘤小鼠的肿瘤细胞提 取其RNA做对照,测定CSF-1及其受体治疗后荷瘤 鼠肿瘤细胞的c-fms、Fra-1及c-Fos等分子的mRNA 表达情况。由RT-PCR电泳结果可见在实验组中 c-fms、Fra-1、c-Fos的mRNA表达量都不同程度地 高于相应对照组。从电泳结果可看出,与对照组相 比实验组小鼠肿瘤细胞中与CSF-1相关的癌基因表 达水平均有不同程度的变化。c-fms、Fra-1及c-Fos 的表达量有所增加,促进了肿瘤增长,见图 3。
 |
图 3RT-PCR检测肿瘤细胞内不同基因的mRNA表达结果 分析 (灰度值,x±s) Figure 3Comparison of mRNA expression in tumor cells between two groups detected by RT-PCR (gray value,x±s) |
为了进一步从数量及功能变化方面了解相关 免疫细胞在荷瘤前后表达的变化,我们采用半定 量RT-PCR技术检测了一些关于免疫抑制相关分 子的mRNA在荷瘤小鼠胸腺、脾脏及淋巴结中的 表达情况,从而推断出相关免疫细胞分泌免疫抑 制性细胞因子的能力。检测的目的分子包括核内 的转录因子Foxp3、表面分子CD152、CD127及分 泌产物TGF-β、IL-10等。从电泳结果可看出,无 论在胸腺、脾脏或淋巴结中,肿瘤组小鼠体内目 的基因表达水平均有不同程度的变化。具体表现 为与对照组相比,实验组CD127表达量下降,而 Foxp3、CD152、TGF-β、IL-10的表达量有所增 加,这表明荷瘤小鼠体内抑制活性的免疫细胞数 量增加,可以推测,在荷瘤小鼠体内,免疫细胞 表现抑制能力增强,CSF-1及其受体治疗后荷瘤鼠 的免疫细胞增值活跃,发挥更强大的免疫抑制功 能,促进了肿瘤增长,见图 4。
 |
图 4RT-PCR检测不同免疫器官中细胞因子及表面分子mRNA表达结果分析(灰度值,x±s) Figure 4Comparison of mRNA expression of cytokines and surface molecules in deferent immune organs between two groups detected by RT-PCR (gray value,x±s) |
CSF-1不仅能选择性刺激巨噬细胞集落形成,诱 导巨噬细胞的增殖分化,而且在许多肿瘤的发生、 发展过程中也扮演重要的角色[5]。通过动物实验检测 CSF-1及其受体对荷瘤鼠肿瘤体积的影响,发现CSF-1 及其受体作用能够加快荷瘤鼠肿瘤的体积增长。
我们便从细胞水平探求原因,发现与正常荷 瘤小鼠相比,实验组荷瘤鼠的肿瘤细胞、脾脏细 胞、骨髓细胞的增殖水平均呈现升高的状态。肿 瘤细胞的增殖活跃是理解范围之内的,由于脾 脏、骨髓部位的淋巴细胞是机体免疫的主要效应 细胞,它的增殖功能在一定程度上反应了机体的 免疫功能状态。因此,促进肿瘤增长结果说明 CSF-1及其受体治疗后荷瘤鼠的整体免疫功能是低 下的。
本实验用RT-PCR分别检测了肿瘤细胞与 CSF-1相关的癌基因表达,结果显示c-fms、 Fra-1、c-Fos的mRNA表达量都不同程度高表达, 这一结果与动物实验结果吻合。RT-PCR的另一 部分结果提示:无论在胸腺、脾脏或是淋巴结 中,肿瘤组小鼠体内的目的基因的表达水平均有 不同程度的变化。其中,CD127表达量下降,而 Foxp3、CD152、TGF-β、IL-10的表达量有所增 加。CD127low低提示抑制性CD4+CD25+调节性T 细胞的分泌产物TGF-β、IL-10的mRNA表达量上 升,而CD4+CD25+调节性T细胞功能的发挥有赖 于这两种细胞因子,同时Foxp3及CD152也影响着 CD4+CD25+调节性T细胞功能的发挥,它们的基因 表达水平上调,表明CD4+CD25+调节性T细胞的功 能增强,导致机体免疫功能低下,免疫系统对肿 瘤免疫应答降低,有利于肿瘤耐受的形成,促进 肿瘤的发生和发展。换言之,CSF-1及其受体治疗 后荷瘤鼠的免疫细胞增殖活跃,发挥更强大的免 疫抑制功能,促进了肿瘤增长。
CSF-1作为炎症介质分子,结合于膜受体, 受体二聚化引发磷酸化,进而激活下游通路,如 Ras、MAPK和PI3K等,使M-CSF效力于肿瘤的侵 袭和生长,在乳腺癌预后不良的病例中也出现了 较高比例的M-CSF和M-CSFR的过表达[6]。总之, 我们确定CSF-1及其受体促进荷瘤鼠的抑制表型 的免疫细胞增殖活跃,发挥更强大的免疫抑制功 能,从而促进了肿瘤增长[7]。
此次试验结果证实CSF-1及其受体会促进乳 腺癌肿瘤的增长,进一步支持了阻断CSF-1/ CSF- 1R(c-fms)则可以抑制肿瘤生长的观点,这也是肿 瘤免疫抑制机制的一部分。通过阻断CSF-1/ CSF- 1R(c-fms)来逆转抑制性免疫细胞诱导的免疫耐 受,为提高抗肿瘤治疗效果开辟了新途径[8]。由此 可以设想用c-fms基因疫苗可以预防并阻止乳腺癌 肿瘤增长,并能够进一步开展针对性肿瘤免疫治 疗的新技术,以原癌基因为肿瘤免疫的靶点构建 基因疫苗,挖掘肿瘤免疫治疗的潜在价值。
参考文献
| [1] | Stanley ER,Berg KL,Einstein DB,et al.Biology and action of colony--stimulating factor-1[J]. Mol Reprod Dev,1997,46(1):4-10. |
| [2] | Chambers SK,Kacinski BM,Ivins CM,et al.Overexpression of epithelial macrophage colony-stimulating factor (CSF-1) and CSF-1 receptor: a poor prognostic factor in epithelial ovarian cancer, contrasted with a protective effect of stromal CSF-1[J]. Clin Cancer Res,1997,3(6):999-1007. |
| [3] | Yang DH, Liu WW, Gu JR. Expressions of IGF-I,IGF-II receptor and CSF-1 receptor of /c-fms oncogene in hepatocellular carcinomas and adjacent non-tumor tissues[J].Di San Jun Yi Da Xue Xue Bao,1992,14(1):100.[杨冬华,刘为纹,顾健人.IGF-II、 IGF-II受体和CSF-1受体/c-fms癌基因产物在肝癌及癌旁肝组 织的表达[J].第三军医大学学报,1992,14(1):100.] |
| [4] | Pollard JW. Role of colony-stimulating factor-1 in reproduction and development[J]. Mol Reprod Dev, 1997 ,46(1):54-60. |
| [5] | Kacinski BM. CSF-1 and its receptor in breast carcinomas and neoplasms of the female reproductive tract[J]. Mol Reprod Dev, 1997,46(1):71-4. |
| [6] | Qian LF, Wu KF, Geng YQ, et al. Biological effect of monoclonal antibodies against CSF1 & CSF 1R on human hepatic cancer cells transplanted in nude mice[J].Zhong Liu,1997,17(4):207-8. [钱连 芳,吴克复,耿以琪,等.集落刺激因子1及其受体的单抗对人肝癌 细胞在裸鼠体内生长的抑制作用[J].肿瘤,1997,17(4):207-8.] |
| [7] | Cui J,Yang DH.CSF-1 receptor/c-fms and liver cancer[J].Shi Jie Hua Ren Xiao Hua Za Zhi,2000,8(6):696-7.[崔俊,杨冬华. CSF-1 受体/c-fms与肝癌[J].世界华人消化杂志,2000,8(6):696-7.] |