 2014, Vol.41
2014, Vol.41


卵巢癌细胞上皮间质转化及侵袭转移过程中microRNA-9的调控作用
引用本文
孙朝阳,李 娜,周 波,杨宗元,卢运萍,翁丹卉. 卵巢癌细胞上皮间质转化及侵袭转移过程中microRNA-9的调控作用[J]. 肿瘤防治研究, 2014, 41(04): 316-319. 
SUN Chaoyang, LI Na, ZHOU Bo,YANG Zongyuan,LU Yunping,WENG Danhui.miR-9 Regulates Epithelial-to-Mesenchymal Transition,Invasion and Metastasis in Ovarian Cancer Cells. Cancer Research on Prevention and Treatment, 2014, 41(04): 316-319.
卵巢癌细胞上皮间质转化及侵袭转移过程中microRNA-9的调控作用
430030 武汉,华中科技大学同济医学院附属同济医院妇产科
摘要:目的 探讨microRNA-9(miR-9)参与调控卵巢癌细胞EMT过程,以及影响卵巢癌细胞侵
袭及转移的分子机制。方法 使用TargetScan及PicTar数据库,预测可能靶向E-cadherin 3’UTR区的
miRNA,双荧光素酶报告体系进行验证;上调候选miR后,用qRT-PCR和Western blot检测E-cadherin
的表达变化;细胞免疫荧光观察E-cadherin、β-Catenin和Vimentin的表达;划痕实验、Transwell实验,
观察卵巢癌细胞运动和侵袭能力的改变。结果 TargetScan、PicTar预测发现miR-9是唯一可与CDH1
结合的miRNA。双荧光素酶报告系统验证预测结果正确。在SKOV-3中上调miR-9后,E-cadherin的表
达受到显著抑制;细胞形态向间质细胞样转变,发生EMT分子水平的特征性改变;体外实验表明,
卵巢癌细胞的运动和侵袭能力得到明显促进。结论 miR-9可以通过靶向调控E-cadherin表达,促进卵
巢癌细胞的EMT进程,对卵巢癌细胞的运动和侵袭能力产生重要影响。
关键词:
microRNA-9
上皮间质转化(EMT)
E-钙粘蛋白
卵巢癌
miR-9 Regulates Epithelial-to-Mesenchymal Transition,Invasion and Metastasis in Ovarian Cancer Cells
Department of Gynecology and Obstetrics, Tongji Hospital Affliated of Tongji College, Tongji
Medical University,Wuhan 430030,China
Abstract:Objective To determine the potential role of miRNAs in controlling EMT and metastasis in
ovarian cancer cells. Methods Firstly, we used 2 computational algorithms, TargetScan and PicTar, to search
for miRNAs that could target 3’ UTR of CDH1 mRNA. Luciferase reporter assay, qRT- PCR and Western blot
were carried out to validate the results. Immunofluorescence staining of E-cadherin, β-Catenin and Vimentin
were performed to detect the EMT process of ovarian cancer cell lines transfected with miRNAs. Wound
healing assay and Transwell assay were used to detect the cell motility and invasiveness. Results miR-9
directly regulated CDH1, causing a remarkable inhibition of E-cadherin expression. miR-9 also caused a
dramatic increasing of ß-Catenin and Vimentin expression, which were the important molecular markers
of EMT process. Moreover, up-regulation of miR-9 in ovarian cancer cell lines significantly improved cell
motility and invasiveness. Conclusion miR-9 has a significantly promoting effect on EMT via directly
targeting E-cadherin in ovarian cancer cells, and may play a potential important role in cancer metastasis and
invasiveness.
Key words:
microRNA-9
Epithelial-to-mesenchymal transition(EMT)
E-cadherin
0 引言
2.2 体外验证miR-9 调控E-cadherin
2.3 上调miR-9表达促进卵巢癌细胞EMT进程
2.4 上调miR-9表达促进卵巢癌细胞运动和侵袭能力
3 讨论
卵巢癌恶性程度高,死亡率位居妇科肿瘤 之首。75%患者确诊时已是晚期,多伴有腹腔等 远处转移,5年生存率约为25% [1]。上皮间质转化 (epithelial-mesenchymal transition,EMT)是肿瘤侵 袭转移中发生的重要特征[2, 3]。在卵巢癌中,EMT 的进程也被证实与肿瘤进展及转移密切相关[4, 5]。 E-cadherin不仅是EMT进程的重要分子标志,同时 作为EMT进程的关键分子,是研究调控肿瘤细胞 EMT机制的重要分子靶标[3, 6, 7]。研究表明E-cadherin 在卵巢癌组织中的表达下调,但是机制仍然不十分 明确。microRNA是大约由20到25个核苷酸构成、进 化上高度保守的一类非编码RNA [8, 9]。它 通 过 碱 基 互 补配对的方式与靶基因的3’UTR区结合,在转录后 水平调控靶基因的表达[10],对于多种肿瘤细胞的生 长、分化、侵袭和转移有着重要的调控作用 [11, 12]。
为了探索miRNA与卵巢癌E-cadherin表达异常 之间的关系,本课题组首先用生物信息学分析了可 能与E-cadherin表达调控有关的miRNA,发现miR-9 是唯一能与E-cadherin编码基因(CDH1)3’UTR 区发生结合的miRNA。由此,本研究对miR-9对 E-cadherin表达的调控作用及其对卵巢癌细胞侵袭 和转移能力的影响进行了探讨。 1 材料和方法 1.1 主要试剂和仪器
RPMI 1640培养液购自美国Gibco公司;miR-9 mimics及所用引物购自中国广州锐博生物科技有 限公司。Dual-luciferase检测试剂盒购自Promega公 司;E-cadherin,Vimentin,β-Catenin抗体购自美 国Epitomics公司;Transwell培养板由美国Corning 公司生产;Matrigel 购自美国BD公司。 1.2 生物信息学miRNA靶基因预测
使用目前公认的miRNA靶基因预测数据库 TargetScan、(http://www.targetscan.org/)、PicTar (http://pictar.mdc-berlin.de/),预测哪些miRNA 可以靶向E-cadherin的编码基因CDH1,并使用 RNA22软件计算两者的结合自由能。 1.3 细胞培养及转染
卵巢癌细胞株SKOV-3购自美国标准生物 品收藏中心(ATCC)。用含有10%胎牛血清的 DMEM培养液,5% CO2条件下传代培养。转染 前取每孔4×105个细胞接种六孔板,培养过夜后, 使用Invitrogen LipofectamineTM 2000转染miR-9 mimics(100 μM) 试剂,转染miR-9 mimics 48 h后进 行后续实验。 1.4 双荧光素酶报告基因体系
构建含有CDH1 mRNA的3’UTR的双荧光素 酶报告基因体系,称为psi-check2-CDH1。miR-9 mimics 分别和psi-check2-CDH1及对照psi-check2共 转染,24 h后使用Dual-luciferase检测试剂盒检测 luciferase活性。 1.5 Western blot检测E-cadherin表达
取转染miR-9 mimics后48 h的SKOV-3细胞,裂 解细胞提取总蛋白。一抗(1:1 000稀释)4℃孵育 过夜,β-actin作为内参照,结果用Image J软件分 析条带灰度值。 1.6 细胞免疫荧光
取转染miR-9 mimics 24 h及对照组的SKOV-3细 胞,消化接种至无菌载玻片上,培养24 h,4%多聚甲 醛固定10 min,PBS冲洗,Triton100破膜5 min,PBS 冲洗,羊血清封闭室温30 min,分别加入E-cadherin (1:50稀释)、Vimentin(1:100稀释)、β-Catenin (1:100稀释)抗体,4℃孵育过夜,PBS冲洗, 加入绿色荧光二抗(1:200稀释)37℃避光孵育30 min,之后均避光操作,PBS冲洗,DIPI工作液染 核5 min,荧光显微镜下观察并拍照。 1.7 Transwell试验(细胞侵袭试验)
Matrigel用DMEM培养液1:5稀释,每孔80 ul加 入Transwell上室,37℃放置1 h备用。24孔板每孔 加入600 μl含10%胎牛血清的RPMI1640培养液,放 入Transwell上室,每小室内加入200 μl无血清培养 液,内含含5×103个细胞。培养24 h后取出上室, 用4%多聚甲醛固定10 min,1 g/L的结晶紫染色10 min,PBS冲洗3次,风干后置于显微镜下观察, 随机计数6个高倍镜视野计数细胞数,拍照并用 ImageJ软件分析。 1.8 统计学方法
采用SPSS 18.0统计软件处理数据,计量资料 用t检验分析。P<0.05为差异有统计学意义。 2 结果 2.1 miR-9可与CDH1 mRNA的3’UTR区域结合
通过目前公认的miRNA靶基因预测数据库 TargetScan和PicTar预测可以靶向E-cadherin编码基 因CDH1的miRNAs。结果发现只有miR-9可以结合 于CDH1 mRNA 3’UTR区的第1327-1333号碱基, 见图 1A。RNA22软件计算结合自由能为-26.40 Kcal/mol,并且生物信息学分析显示和miR-9种子 序列特异性结合的CDH1 3’UTR序列存在高度种属 保守性,见图 1B。
 | 图 1 CDH1 mRNA的3’UTR区与miR-9相结合 Figure 1 miR-9 directly targets CDH1 by interacting with its 3’UTR A: the binding site of CDH1 mRNA 3’UTR predicted by TargetScan and PicTar algorithms analysis; B:the predicted binding site of CDH1 mRNA 3’UTR with miR-9 was highly conserved in many species, and the complementary sequences of seed sequence of miR-222 were highlighted |
构建含有CDH1 mRNA 3’UTR的双荧光素酶报 告载体,称为psi-check2-CDH1。miR-9 mimics 分 别和psi-check2-CDH1及对照psi-check2共转染,发 现miR-9可以降低psi-check2-CDH1组海肾荧光素 酶活性到58%(t=7.937,P=0.001),见图2A,提示 miR-9可以与CDH1的3’UTR区相结合。
在SKOV-3中,转染miR-9 mimics 48 h,Realtime PCR检测发现miR-9表达上调为对照组的23.4 倍(t=8.257,P=0.001);CDH1 mRNA表达降低 至对照组的24%(t=16.8,P=0.000),见图2B; Wester n blot法检测E-cadherin的表达明显下降,为 对照组的26%(t=28.3,P=0.000),见图2C。
 | 图2 体外验证CDH1为miR-9的靶基因
Figure 2 Validation that miR-9 directly targets CDH-1 in vitro
A:Luciferase activities were measured in SKOV3 cells cotransfected with
psi-check2-CDH1 with miR-9 mimic or miR-control; B: Real-time PCR
of miR-9 and CDH1 mRNA in miR-9 mimic or miR control transfected
cells; C: Immunoblotting of E-cadherin in miR-9 mimic or miR-control
transfected cells. Values represented 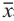 ±s,n=3;*:P<0.05 ±s,n=3;*:P<0.05
|
转染miR-9 mimics 48 h后,显微镜见SKOV-3-miR-9细胞间连接更为松散,胞质向外延展成“伪 足”样,表现为成纤维细胞或间质细胞形态,见图 3A。免疫荧光结果显示,与对照组相比,SKOV-3-miR-9组上皮性标志物E-cadherin在胞质内表达明 显下降,而间质性标志物Vimentin和β-Catenin的荧 光强度显著增强,见图3B。
 | 图3 上调miR-9促进卵巢癌细胞系SKOV-3的EMT进程 Figure 3 miR-9 upregulation promotes EMT transformation of SKOV-3 cell A:phase contrast images illustrated that miR-9 mimic transfected SKOV-3 cells showed EMT morphological changes;B: Immunofluorescence staining of E-cadherin,Vimentin,β-Catenin (green) in miR-9 mimic or miR control transfected cells |
划痕实验发现SKOV-3-miR-9划痕愈合程度明 显快于SKOV-3,使用TScratch软件分析划痕面细 胞覆盖面积,48 h细胞覆盖面积为93%,对照组为 61%,两者差异有统计学意义(t=3.215,P=0.032), 见图4A。Transwell试验显示,SKOV-3-miR-9细 胞在24 h后穿过上室的细胞均数为120.33个/孔, 对照组为6个/孔,差异有统计学意义(t=13.527, P=0.000),见图4B。
 | 图4 上调miR-9促进卵巢癌细胞的运动和侵袭能力 Figure 4 miR-9 upregulation promotes migration and invasion of SKOV-3 cell A: wound healing test of miR-9 mimic or miR control transfected SKOV-3 cells;B: transwell invasion assay of miR-9 mimic or miR control transfected SKOV-3 cells; *: P<0.05 |
在包括卵巢癌在内的很多肿瘤组织,尤其是 在转移灶中,都发现E-cadherin的异常表达,提示 E-cadherin的表达情况与肿瘤转移及远期预后密切 相关[13, 14, 15]。作为EMT的关键分子,E-cadherin异常 表达可能受多种机制调控,如启动子甲基化、基 因突变等,还有研究表明miR-200家族等miRNA也 参与了EMT过程中E-cadherin的调控[16, 17]。因此针 对靶向E-cadherin的miRNA的深入探索,对于揭示 肿瘤转移的分子机制和研究抗肿瘤转移药物靶点 有重要意义。本文证明了miR-9可以通过负向调控 E-cadherin表达,诱导卵巢癌细胞的EMT进程,进 而促进卵巢癌细胞的运动和侵袭能力,并可以进 一步推测,miR-9的表达高低对卵巢癌的转移和预 后具有重要影响。这为卵巢癌EMT过程的信号通 路和分子机制的研究提供了依据,为如何抑制癌 细胞转移的卵巢癌靶向治疗提供了新的思路。
miR-9在多种肿瘤中被证实为肿瘤相关miRNA, 可以影响不同的肿瘤细胞的生长、转移或耐药 等。Iorio等[18]研究发现,与癌旁的正常乳腺组织 相比,乳腺癌组织中存在miR-9的高表达;在人 神经胶质瘤中,也报道高表达的miR-9通过调控 Chromobox 7(CBX7)参与胶质瘤的发生发展 [19]。 然而对miR-9的表达水平与肿瘤的关系,研究者 们也有不一致的看法。Laios等[20]发现,miR-9的 低表达能通过通过调控bcl-2,CLDN等诱导卵巢 癌的化疗耐药。由于miRNA和其靶基因作用关系 网络复杂,一个miRNA可以调控多个靶基因,一 个靶基因可以被多个miRNA调控,形成复杂的反 馈调节关系,因此推测miR-9不仅可以通过调控 E-cadherin进而诱导卵巢癌细胞EMT进程,促进侵 袭转移能力,还有可能通过其他的作用靶点来影 响卵巢癌细胞的生长、耐药等其他恶性生物学行 为。如何更全面地阐明肿瘤相关miRNA在卵巢癌 发生发展中的作用,仍需进一步探索。
参考文献
| [1] | Colombo N, Van Gorp T, Parma G, et al. Ovarian cancer[J]. Crit Rev Oncol Hematol, 2006, 60(2):159-79. |
| [2] | López-Novoa JM, Nieto MA. Inflammation and EMT: an alliance towards organ fibrosis and cancer progression[J]. EMBO Mol Med, 2009, 1(6-7):303-14. |
| [3] | Thiery JP. Epithelial-mesenchymal transitions in tumour progression[J]. Nat Rev Cancer, 2002,2(6):442-54. |
| [4] | Ahmed N, Thompson EW, Quinn MA. Epithelial-mesenchymal interconversions in normal ovarian surface epithelium and ovarian carcinomas: an exception to the norm[J]. J Cell Physiol, 2007, 213(3):581-8. |
| [5] | Cipollone JA, Graves ML, K?bel M, et al. The anti-adhesive mucin podocalyxin may help initiate the transperitoneal metastasis of high grade serous ovarian carcinoma[J]. Clin Exp Metastasis, 2012, 29(3):239-52. |
| [6] | Shah PP, Kakar SS. Pituitary tumor transforming gene induces epithelial to mesenchymal transition by regulation of Twist, Snail, Slug, and E-cadherin[J]. Cancer Lett, 2011, 311(1):66-76. |
| [7] | Yu L, Li HZ, Lu SM, et al. Down-regulation of TWIST decreases migration and invasion of laryngeal carcinoma Hep-2 cells by regulating the E-cadherin, N-cadherin expression[J]. J Cancer Res Clin Oncol, 2011, 137(10):1487-93. |
| [8] | Davis-Dusenbery BN, Hata A. MicroRNA in cancer: the involvement of aberrant MicroRNA biogenesis regulatory pathways[J]. Genes Cancer, 2010,1(11):1100-14. |
| [9] | Gregory RI, Shiekhattar R. MicroRNA biogenesis and cancer[J]. Cancer Res, 2005, 65(9):3509-12. |
| [10] | Valencia-Sanchez MA, Liu J, Hannon GJ, et al. Control of translation and mRNA degradation by miRNAs and siRNAs[J]. Genes Dev, 2006, 20(5):515-24. |
| [11] | Wang HB, Zhang HJ, Su JM,et al. Epithelial-mesenchymal transition (EMT) and its effect on microRNA expression in lung cancer[J]. Zhonghua Zhong Liu Za Zhi, 2011, 33(8):590-3.[王海兵,张慧君,苏晋梅,等. 肺癌A549细胞上皮-间质转化及其对微小RNA表达的影响[J].中华肿瘤杂志, 2011, 33(8):590-3.] |
| [12] | Bryant JL, Britson J, Balko JM, et al. A microRNA gene expression signature predicts response to erlotinib in epithelial cancer cell lines and targets EMT[J]. Br J Cancer, 2012, 106(1):148-56. |
| [13] | Ho CM, Cheng WF, Lin MC, et al. Prognostic and predictive values of E-cadherin for patients of ovarian clear cell adenocarcinoma[J]. Int J Gynecol Cancer, 2010, 20(9):1490-7. |
| [14] | Huang KJ, Sui LH. The relevance and role of vascular endothelial growth factor C, matrix metalloproteinase-2 and E-cadherin in epithelial ovarian cancer[J]. Med Oncol, 2012, 29(1):318-23. |
| [15] | Koensgen D, Freitag C, Klaman I, et al. Expression and localization of e-cadherin in epithelial ovarian cancer[J]. Anticancer Res, 2010,30(7):2525-30. |
| [16] | Li Y, VandenBoom TG 2nd, Kong D, et al. Up-regulation of miR-200 and let-7 by natural agents leads to the reversal of epithelial-to-mesenchymal transition in gemcitabine-resistant pancreatic cancer cells[J]. Cancer Res, 2009, 69(16):6704-12. |
| [17] | Schickel R, Park SM, Murmann AE,et al. miR-200c regulates induction of apoptosis through CD95 by targeting FAP-1[J]. Mol Cell, 2010, 38(6):908-15. |
| [18] | Iorio MV, Ferracin M, Liu CG, et al. MicroRNA gene expression deregulation in human breast cancer[J]. Cancer Res, 2005, 65(16):7065-70. |
| [19] | Chao TF, Zhang Y, Yan XQ,et al. MiR-9 regulates the expression of CBX7 in human glioma [J]. Zhongguo Yi Xue Ke Xue Yuan Xue Bao, 2008, 30(3):268-74.[晁腾飞,张瑜,颜兴起,等. MiR-9在人神经胶质瘤中调节CBX7的表达[J].中国医学科学院学报, 2008, 30(3):268-74.] |
| [20] | Laios A, O’Toole S, Flavin R, et al. Potential role of miR-9 and miR-223 in recurrent ovarian cancer[J]. Mol Cancer, 2008, 7:35. |