
Notch Signaling Pathway Regulates Epithelial-mesenchymal Transition of Osteosarcoma by Activating Slug
-
摘要:目的
探究Notch信号通路对骨肉瘤上皮-间质转化的影响及其机制。
方法通过慢病毒转染技术转染骨肉瘤细胞, 利用si-slug敲除Slug基因, 采用免疫荧光染色技术、Western blot、qRT-PCR、相差显微镜、划痕实验和TranswellTM实验检测相应骨肉瘤细胞形态、侵袭、转移及上皮间质转化(EMT)情况。动物实验比较各组小鼠成瘤后体积、重量变化; 采用qRT-PCR检测离体肿瘤组织中Slug和N-cadherin的表达。
结果成功得到Notch信号通路激活或抑制的骨肉瘤细胞, Notch信号通路激活组E-cadherin表达无明显变化, N-cadherin表达显著升高, 骨肉瘤细胞长/短轴比率上调, Slug、Twist1表达上调, 侵袭及转移能力增强。动物实验发现Notch信号通路激活可促进体内成瘤, 离体骨肉瘤组织中Slug、N-cadherin阳性率增高。Slug被成功敲除后Notch信号激活组骨肉瘤细胞形态、侵袭及转移能力发生逆转。
结论Notch信号通路对骨肉瘤上皮-间质转化过程具有促进作用, 其机制可能与激活Slug有关。
Abstract:ObjectiveTo investigate the effect and mechanism of Notch signaling pathway on epithelial-mesenchymal transition (EMT) of osteosarcoma.
MethodsWe used lentivirus transfection technology to transfect osteosarcoma cells, and used si-slug to knock down the Slug gene. Then we used immunofluorescence staining, Western blot, qRT-PCR, phase contrast microscopy, wound healing and TranswellTM test to detect the morphological, the invasion and metastasis and the influences of EMT of the corresponding osteosarcoma cells. In animal experiments, the volume and weight of nude mice were compared after tumorigenesis, and the expression of Slug and N-cadherin in tumor tissues were detected by qRT-PCR.
ResultsWe successfully obtained osteosarcoma cells with activated or inhibited Notch signaling pathway. In Notch activated group, the expression of N-cadherin was increased significantly, the ratio of long/short axis, the expression of Slug and Twist1, as well as the invasion and metastasis abilities of osteosarcoma cells were increased. Animal experiments showed that activated Notch signaling pathway could promote tumorigenesis in vivo, and the positive rates of Slug and N-cadherin in osteosarcoma tissues in xenograft were upregulated. The morphology, invasion and metastasis of osteosarcoma cells in the Notch activated group were reversed after Slug was successfully knocked out.
ConclusionNotch signaling pathway promotes the EMT of osteosarcoma, which may be related to Slug activation.
-
Key words:
- Osteosarcoma /
- EMT /
- Notch signaling pathway /
- Slug
-
0 引言
肺癌是全球范围内发病率和死亡率最高的恶性肿瘤。2017年死于肺癌的总人数超过乳腺癌、前列腺癌、结直肠癌和脑瘤的总和,预计2020年死于肺癌的人数将达到所有癌症死亡人数的22%以上[1],局部复发和远处转移是导致肺癌高病死率的主要原因。肿瘤的异质性是单次活检无法获取肿瘤全部分子信息的重要原因,也是造成肿瘤耐药性的基础,因此需要对肿瘤的分子分型进行动态评估。液体活检作为一种非侵入性的肿瘤检测方法,可持续、动态地评估肿瘤细胞的基因突变状态[3]。循环肿瘤细胞(circulating tumor cell, CTCs)、循环肿瘤DNA(circulating tumor DNA, ctDNA)、外泌体、mi-RNA、cf-DNA等是肺癌液态活检的主要检测指标[4],其中CTCs可以提供包含DNA、RNA和蛋白质等多层次的分子信息,而其他指标只能提供遗传层面的异常信息[5]。可见CTCs在肺癌的诊断、疗效评价、预后预测等方面作用显著[6]。本文讨论了CTCs的转移级联过程、检测方法,以及肺癌CTCs在临床早期筛查、预测复发转移、疗效监测评价等方面的应用,并融合入中医“正虚伏毒”理论,以期为肺癌转移转化研究提供新的思路。
1 肺癌循环肿瘤细胞
1869年,Ashworth首次提出循环肿瘤细胞的概念,CTCs是从原发肿瘤脱落进入血液的肿瘤细胞,从浸润至进入循环会发生一系列转移级联过程进而造成远处转移和定植[7]。2019年,田建辉等从一例Ⅱa期肺癌患者的血液中分离出CTCs,成功建立了永久性的人肺腺癌循环肿瘤细胞系CTC-TJH-01。CTC-TJH-01处于上皮间质转化(EMT)的中间阶段,具有干细胞表型、耐药性强和休眠的特性,在NOD-SCID小鼠上具有致瘤性和肺转移能力[8]。根据EMT的不同阶段可将CTCs亚群分为上皮-间充质型循环肿瘤细胞、上皮型循环肿瘤细胞和间充质型循环肿瘤细胞[9],不同表型的CTCs转移能力有所差异。
2 CTCs参与肿瘤转移级联过程
2.1 肿瘤细胞脱落
肿瘤细胞从原发灶脱落浸润基底膜主要有两种方式:(1)EMT介导途径。EMT是上皮细胞向间质细胞转化的关键过程,通过分泌相关转录因子(Snail、Twist、ZEB-1)、细胞外分子(TGF-β、HGF)、炎性细胞以及特定的信号转导途径(Wnt、β-catenin、NF-κB、PI3K)调节相关蛋白(上皮标志物:EpCAM、E-cadherin、CK;间质标志物:Vimentin、N-cadherin等)表达,使肿瘤细胞向间充质型转化,从而获得更强的移动性并抵抗细胞凋亡[10];(2)非EMT介导途径。肿瘤细胞通过外力和中心体增大的方式,介导肿瘤细胞被动渗入血液,以单细胞或细胞簇的形式离开原发肿瘤[11]。通过以上两种方式进入循环的CTCs可能具有不同的细胞表型,皆可引起肿瘤转移。
2.2 血液循环中的CTCs
实体肿瘤每天都有数以万计的肿瘤细胞进入血液循环,CTCs在血液循环中面临着免疫监视、血流剪切力、氧化应激等多方面的因素,只有未被免疫系统清除的CTCs才能进一步播散定植[12]。其中单个循环肿瘤细胞易被免疫细胞清除,发生失巢凋亡。而由多个循环肿瘤细胞组成的循环肿瘤细胞簇或循环肿瘤微栓(circulating tumor microemboli, CTM)能更大程度上发生免疫逃逸,具有更强的凋亡抵抗,更易发生转移[13]。血液循环中的CTCs可被血小板识别,激活血小板并诱导相关细胞因子释放和聚集,形成由血小板包被的细胞簇,从而逃避NK细胞和T细胞的杀伤作用,进而发生免疫逃逸[14]。
2.3 CTCs远处转移和定植
进入血液循环的CTCs可以锚定在内皮细胞的表面,通过受体-配体结合的方式使CTC发生外渗,进而产生远处转移。不同的是,单个CTCs通过毛细血管向外迁移,而CTC簇可逆性地重组为单排链,再横向穿过毛细血管[15]。CTCs表面的整联蛋白通过与内皮细胞表面的黏附分子1(ICAM1)或血管细胞黏附分子1(VCAM1)结合,为CTCs突破血管壁细胞外基质提供动力。CTCs产生的趋化因子配体取决于各自受体的表达,进而决定远处的归巢器官[16]。CTCs还可调节肿瘤微环境中相关抑制性免疫细胞以支持自身增殖和存活。
3 肺癌循环肿瘤细胞检测
CTCs的富集和分离主要有两种方式:其一,基于CTCs独特的物理特性,通过过滤或密度梯度离心等方式获取,被称为“非标记依赖法”;其二,基于CTCs经历的EMT过程,表达特异性细胞表面蛋白,可通过免疫磁珠分选、抗体特异性结合等方式获得,被称为“标记依赖法”,最常用的细胞表面标志物是上皮细胞黏附分子(EpCAM)、细胞角蛋白(CK)等。目前常用的基于物理方法的CTCs检测系统主要有:ISET、MetaCell、CellSieve、Parsortix、OncoQuick、VitaAssay等;基于标记依赖的CTCs检测仪器主要有:CellSearch、MagSweeper、MACS、EPISPOT、CellCollector等[17]。然而,不同系统的CTCs检出效率有差异,如Tamminga等比较了CellSearch和ISET对肺癌CTCs的检出率,发现CellSearch的检出率为69%,而ISET的检出率为88%,表明ISET更适合于肺癌患者的CTCs检测[18]。
研究者正在对检测系统进行优化研究,研发出多种不同原理的CTCs检测方法和新型检测仪器,以提高肺癌CTCs的检出率:(1)微流控装置。Wang等设计的集成微流控装置对肺癌CTCs的平均回收率达到92.5%,纯度达到94%[19];(2)芯片法。Yan等则使用集成多功能电化学芯片将CTCs的捕获效率控制在8%~100%[20];(3)免疫磁珠分离法。通常采用上皮细胞黏附分子(EpCAM)作为主要标志物,将与磁性微粒结合的EpCAM抗体加入外周血中,通过免疫磁珠分离富集CTCs[21];(4)纳米技术。核酸适配体修饰的PEG-PLGA-纳米纤维(PPN)微流控系统,可识别肺癌患者中罕见的CTC亚型并进行优化,显著提高对多克隆源性肿瘤细胞的捕获效率[22];(5)新型肺癌CTCs检测仪器。光流式细胞仪(OFCM)集成了多级微流控芯片和四色荧光检测系统,可以完成自动肺癌CTCs分离,进行3D聚焦和单细胞表型分析[23]。
4 肺癌循环肿瘤细胞的临床应用
4.1 早期筛查
肺癌患者在癌症发展早期基本处于无症状阶段,多数患者明确诊断时已至中晚期,已发生转移,错失治疗的最佳时机。肺癌的早期筛查主要通过低剂量计算机断层扫描(LDCT)实现,然而LDCT具有高假阳性,令患有非恶性结节的患者面临不必要的活检相关风险[24]。Duan等使用CellCollector检测了44名肺结节患者和20例健康志愿者外周血中的CTCs,在志愿者外周血中未检测到CTCs。通过病理学诊断结果对比后发现,肺结节患者外周血中CTCs阳性率为52.94%,其诊断特异性为90%,CellCollector系统检测CTCs可有效区分良恶性结节,用于肺癌早期筛查[25]。Li等采用阴性富集-荧光原位杂交(NE-FISH)方法检测肺癌患者及健康志愿者外周血中CTCs水平,发现肺癌患者的CTCs数量明显高于志愿者或患有良性肺疾病的患者,将CTCs与血清肿瘤标志物联合检测可提高早期肺癌患者的诊断敏感度[26]。
4.2 预后及复发转移预测
肺癌的预后与分期密切相关。大量研究表明,检测肺癌患者手术、放化疗前后血液中的CTCs数量和亚群可预测预后,指导进一步的治疗。Wang等发现放射治疗后NSCLC患者外周血中PD-L1+CTC亚群比例升高,预示着治疗反应与预后不良[27]。Tamminga等研究发现晚期NSCLC患者体内CTCs的存在与酪氨酸激酶抑制剂(TKI)或化疗反应率低、无进展生存期(PFS)和总生存期(OS)短相关[28]。复发转移是导致肺癌患者死亡、生存期短和生活质量低的主要原因。患者术后伴有原发灶切除和淋巴结清扫,短期内无法评估和发现复发转移等情况。有研究发现,手术可以导致肺癌患者术后CTCs数目减少,但术后1天与3天的CTCs数目出现早期反弹现象,且与几月后的复发有关[29]。Chinniah等对48例接受放化疗的局部晚期非小细胞肺癌(LA-NSCLC)患者的CTCs进行纵向监测发现,在患者外周血中检测到CTCs早于放射影像学证据显示疾病复发[30]。
4.3 化疗及免疫靶向治疗的疗效监测与评价
由于肿瘤细胞的异质性,存在基因突变(如ALK重排、EGFR突变等)的患者易发生耐药突变,此时需要进行重复的基因检测。通过对捕获的CTCs进行单细胞测序可以检测到EGFR、MET、ALK和ROS1等基因的分子变化以及PD-L1的表达,鉴定肺癌患者免疫靶向治疗后的耐药性突变,其动态变化有助于实时疗效监测及个性化治疗方案的选择[31]。Guibert等研究发现,NSCLC患者外周血中分离的CTCs表面PD-L1阳性率(83%)高于癌组织(41%),且在使用PD-L1抑制剂治疗的患者中,治疗前PD-L1+CTCs与预后不良相关[32]。
5 创新肺癌亚临床研究思路,提出并验证“正虚伏毒”学说
早期肺癌术后患者影像学评估无可见病灶,临床评估为治愈,但是其外周循环和器官组织中还存有CTCs或休眠的肿瘤细胞,患者实际上处于肺癌转移的亚临床状态,这将成为日后复发转移发生的根源。针对该阶段病理特征,田建辉传承《道德经》“有生于无”和《黄帝内经》重视“正气”思想,融合现代肿瘤学和免疫学进展,提出“正虚伏毒”学说并系统验证,“正虚”主要免疫衰老和免疫逃逸为主的免疫抑制内环境,“伏毒”指高危人群或术后患者体内存在的肿瘤干细胞、循环肿瘤细胞和休眠肿瘤细胞等,“伏毒”具有“毒自内生,深伏血道,内藏脏腑,流注全身,伺机为患,正盛则伏而不出,正虚则出而为病”的特点。认为肺癌术后患者存在伏毒是绝对事件,而最终是否发生复发转移与机体的免疫状态密切相关,免疫功能紊乱可以表现为免疫监视和免疫清除能力下降(正虚),促使潜伏在患者循环系统或者脏器内的CTCs(伏毒)、休眠肿瘤细胞和微小转移灶等发生免疫逃逸,最终导致复发与转移的发生[33]。该学说为肺癌复发转移的研究提供了启发和探索。
课题组前期采用阴性富集和阳性富集(微流控芯片结合免疫磁珠分离技术)研究发现肺癌患者外周血中CTC计数与临床分期之间存在负相关性,早期患者外周血中具有较高数目的CTCs。同时研究发现免疫负调控细胞Tregs和MDSCs的比例与肺癌患者的临床分期呈正相关[34]。提示在局部脏器中免疫抑制微环境的形成可能是促进CTCs定植后存活、增殖形成转移灶的原因之一,目前正在进一步研究。此外,田建辉课题组围绕前期建立的肺癌循环肿瘤细胞系(CTC-TJH-01),证实其具有干细胞表型、休眠、免疫逃逸特征,体内研究证实其致瘤性和肺转移特征,目前发现CTCs形成细胞簇后较单个CTCs具有更强的转移能力;而调控NK细胞功能可抑制CTCs肺转移的发生等现象,从而构建了肺癌转移的体内外研究平台,促进肺癌转移转化研究效率的提高[35]。
6 小结与展望
复发转移是导致早期肺癌术后患者死亡的主要因素,也是制约肺癌整体防控效率提高的关键,但是目前仍缺乏成熟的亚临床阶段发病学理论,而循环肿瘤细胞的研究丰富了对该阶段的认识。2018版CSCO《原发性肺癌诊疗指南》肯定了肺癌术后循环肿瘤细胞检测在预测复发中的价值,并逐渐进入临床研究。然而现阶段肺癌CTCs的研究仍局限于检测技术的开发与完善,由于CTCs检测技术具有多样性,而应用不同原理研发的检测系统灵敏度存在差异,导致临床上尚缺乏统一的检测方法和判定标准;另外由于不同CTCs亚群的转移能力也有所区别,仅以CTCs数目预测转移仍存在争议;CTCs最终是否转移与其所处环境因素密切相关,如CTCs与自然杀伤细胞、巨噬细胞等固有免疫细胞;T、B淋巴细胞等适应性免疫细胞以及血小板等存在复杂的作用机制,有待进一步研究探索[36]。因此,通过精准鉴定转移潜能强的CTCs亚群,进而与免疫、凝血功能状态结合构建肺癌转移风险模型是促进临床推广的关键。充分发挥中医学认知疾病的系统性优势,融合现代科学精华不断创新肺癌转移理论和研究体系,将促进突破制约转移疗效提高的瓶颈,推动肺癌整体防控效率的提高。
作者贡献陈敬腾:实验设计与实施、文章执笔余铃:实验设计、文章审校方硕、郭良煜:实验实施郭卫春:实验评估及指导 -
![]()
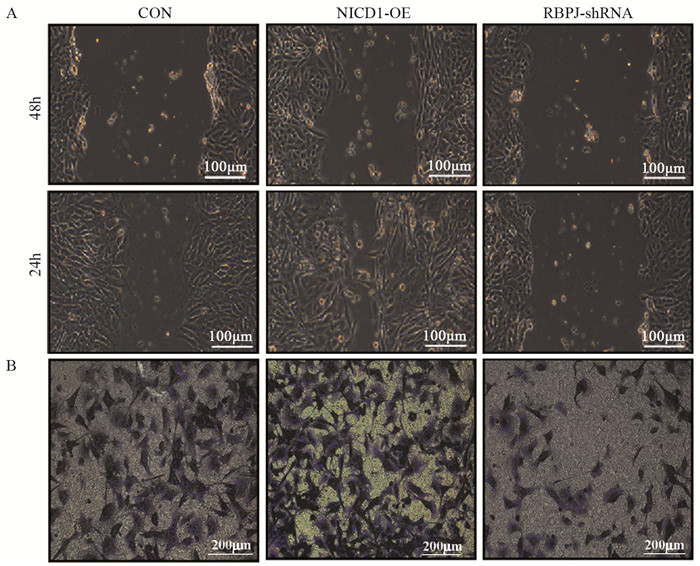
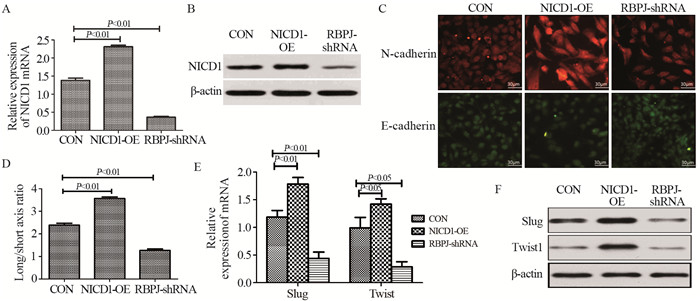
图 1 Notch通路促进骨肉瘤细胞上皮-间质转化
Figure 1 Notch pathway promoted epithelial-mesenchymal transition of osteosarcoma cells
![]()
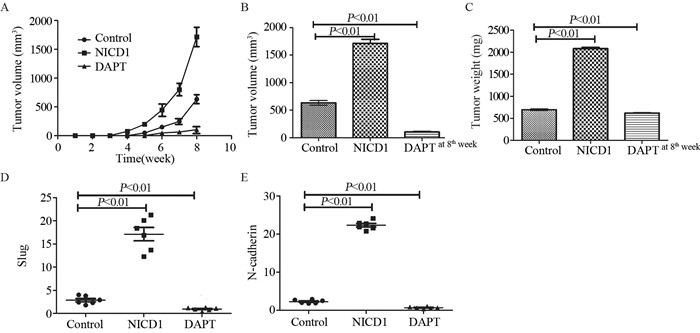
图 2 Notch通路促进骨肉瘤细胞迁移和侵袭
Figure 2 Notch pathway promoted migration(A) and invasion(B) of osteosarcoma cells
![]()
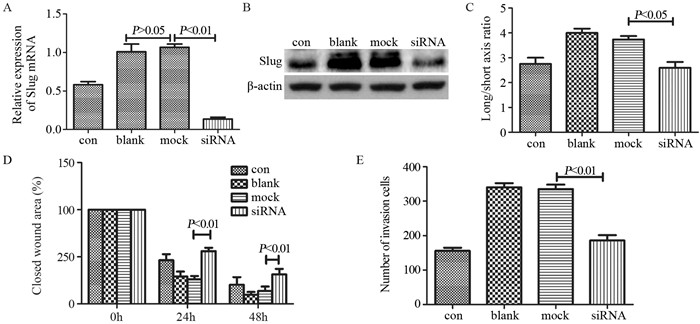
图 3 Notch通路促进骨肉瘤细胞体内成瘤
Figure 3 Notch pathway promoted tumorigenesis in vivo of osteosarcoma cells
-
[1] Isakoff MS, Bielack SS, Meltzer P, et al. Osteosarcoma:Current Treatment and a Collaborative Pathway to Success[J]. J Clin Oncol, 2015, 33(27):3029-3035. doi: 10.1200/JCO.2014.59.4895
[2] 王勃霖, 姜晓玲, 卢斌, 等.骨肉瘤中miR-17-92基因簇作用的研究进展[J].肿瘤防治研究, 2019, 46(7):635-638. doi: 10.3971/j.issn.1000-8578.2019.18.1988 Wang BF, Jiang XL, Lu B, et al. Research Progress in Effect of miR-17-92 Cluster in of Osteosarcoma[J]. Zhong Liu Fang Zhi Yan Jiu, 2019, 46(7):635-638. doi: 10.3971/j.issn.1000-8578.2019.18.1988
[3] 李华杰, 宗治贤, 陈祝明, 等.骨肉瘤的保肢治疗研究进展[J].系统医学, 2019, 4(4):196-198. http://d.old.wanfangdata.com.cn/Periodical/xtyx201904070 Li HJ, Zong ZX, Chen ZM, et al. Research Progress in Conservative Treatment of Osteosarcoma[J]. Xi Tong Yi Xue, 2019, 4(4):196-198. http://d.old.wanfangdata.com.cn/Periodical/xtyx201904070
[4] Simpson S, Dunning MD, de Brot S, et al. Comparative review of human and canine osteosarcoma:morphology, epidemiology, prognosis, treatment and genetics[J]. Acta Vet Scand, 2017, 59(1):71. doi: 10.1186/s13028-017-0341-9
[5] Persa OD, Niessen CM. Epithelial polarity limits EMT[J]. Nat Cell Biol, 2019, 21(3):299-300. doi: 10.1038/s41556-019-0284-7
[6] Owusu-Akyaw A, Krishnamoorthy K, Goldsmith LT, et al. The role of mesenchymal-epithelial transition in endometrial function[J]. Hum Reprod Update, 2019, 25(1):114-133. doi: 10.1093/humupd/dmy035
[7] Nakaya Y, Sheng G. EMT in developmental morphogenesis[J]. Cancer Lett, 2013, 341(1):9-15. doi: 10.1016/j.canlet.2013.02.037
[8] Wushou A, Hou J, Zhao YJ, et al. Twist-1 up-regulation in carcinoma correlates to poor survival[J]. Int J Mol Sci, 2014, 15(12):21621-21630. doi: 10.3390/ijms151221621
[9] Zhang M, Wang D, Zhu T, et al. RASSF4 Overexpression Inhibits the Proliferation, Invasion, EMT, and Wnt Signaling Pathway in Osteosarcoma Cells[J]. Oncol Res, 2017, 25(1):83-91. doi: 10.3727/096504016X14719078133447
[10] Aster JC, Pear WS, Blacklow SC. The Varied Roles of Notch in Cancer[J]. Annu Rev Pathol, 2017, 12:245-275. doi: 10.1146/annurev-pathol-052016-100127
[11] Ito T, Kudoh S, Ichimura T, et al. Small cell lung cancer, an epithelial to mesenchymal transition (EMT)-like cancer:significance of inactive Notch signaling and expression of achaetescute complex homologue 1[J]. Hum Cell, 2017, 30(1):1-10. doi: 10.1007%2Fs13577-016-0149-3
[12] Zhang J, Kuang Y, Wang Y, et al. Notch-4 silencing inhibits prostate cancer growth and EMT via the NF-kappaB pathway[J]. Apoptosis, 2017, 22(6):877-884. doi: 10.1007/s10495-017-1368-0
[13] De Francesco EM, Maggiolini M, Musti AM. Crosstalk between Notch, HIF-1alpha and GPER in Breast Cancer EMT[J]. Int J Mol Sci, 2018, 19(7). pii:E2011. doi: 10.3390/ijms19072011
[14] Yang J, Guo W, Wang L, et al. Notch signaling is important for epithelial-mesenchymal transition induced by low concentrations of doxorubicin in osteosarcoma cell lines[J]. Oncol Lett, 2017, 13(4):2260-2268. doi: 10.3892/ol.2017.5708
[15] 胡涂, 杨庆诚, 程冬冬.骨肉瘤转移侵袭机制研究进展[J].中国骨与关节杂志, 2015, 1:41-44. doi: 10.3969/j.issn.2095-252X.2015.01.011 Hu T, Yang QC, Cheng DD. Research progress on the mechanism of metastasis and invasion of osteosarcoma[J]. Zhongguo Gu Yu Guan Jie Za Zhi, 2015, 1:41-44. doi: 10.3969/j.issn.2095-252X.2015.01.011
[16] Gou Y, Zhai F, Zhang L, et al. RUNX3 regulates hepatocellular carcinoma cell metastasis via targeting miR-186/E-cadherin/EMT pathway[J]. Oncotarget, 2017, 8(37):61475-61486.
[17] Sharili AS, Allen S, Smith K, et al. Snail2 promotes osteosarcoma cell motility through remodelling of the actin cytoskeleton and regulates tumor development[J]. Cancer Lett, 2013, 333(2):170-179. doi: 10.1016/j.canlet.2013.01.027
[18] Yu Z, Zhang Y, Gao N, et al. Overexpression of miR-506 inhibits growth of osteosarcoma through Snail2[J]. Am J Transl Res, 2015, 7(12):2716-2723. http://cn.bing.com/academic/profile?id=99c0e06837f8beeed26d8c5aa954d5dd&encoded=0&v=paper_preview&mkt=zh-cn
[19] Goossens S, Vandamme N, Van Vlierberghe P, et al. EMT transcription factors in cancer development re-evaluated:Beyond EMT and MET[J]. Biochim Biophys Acta Rev Cancer, 2017, 1868(2):584-591. doi: 10.1016/j.bbcan.2017.06.006
[20] McManus MM, Weiss KR, Hughes DP. Understanding the role of Notch in osteosarcoma[J]. Adv Exp Med Biol, 2014, 804:67-92. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=b60238ba88702033dfb31c5efdf580cb
[21] Saad S, Stanners SR, Yong R, et al. Notch mediated epithelial to mesenchymal transformation is associated with increased expression of the Snail transcription factor[J]. Int J Biochem Cell Biol, 2010, 42(7):1115-1122. doi: 10.1016/j.biocel.2010.03.016
[22] Fang S, Yu L, Mei H, et al. Cisplatin promotes mesenchymallike characteristics in osteosarcoma through Snail[J]. Oncol Lett, 2016, 12(6):5007-5014. doi: 10.3892/ol.2016.5342
 下载:
下载:
计量
- 文章访问数: 1663
- HTML全文浏览量: 462
- PDF下载量: 593

