
Effect of Cyclin G1 on Radiosensitivity of Hepatocellular Carcinoma HepG2 Cells and Its Mechanism
-
摘要:目的
探讨Cyclin G1对肝癌HepG2细胞放射敏感度的影响及其可能的分子机制。
方法通过转染Cyclin G1-siRNA构建HepG2-Cyclin G1-siRNA细胞,通过成克隆分析观察Cyclin G1对肝癌细胞放射敏感度的影响,流式细胞仪检测肝癌细胞细胞周期分布,ELISA法检测乏氧因子HIF-1α及ROS的表达,Western blot检测凋亡相关蛋白Bcl-2、Bax的表达。
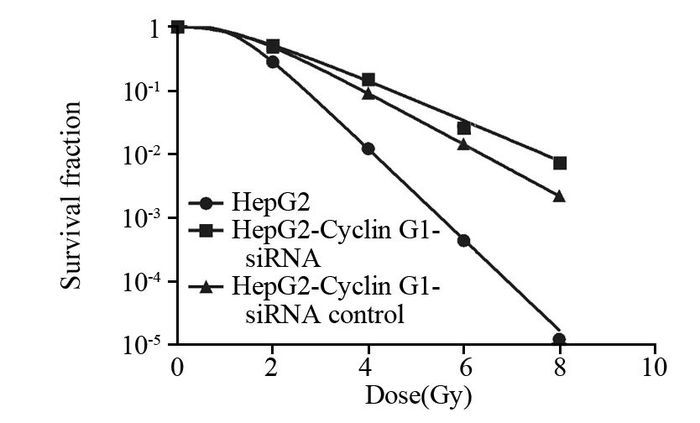
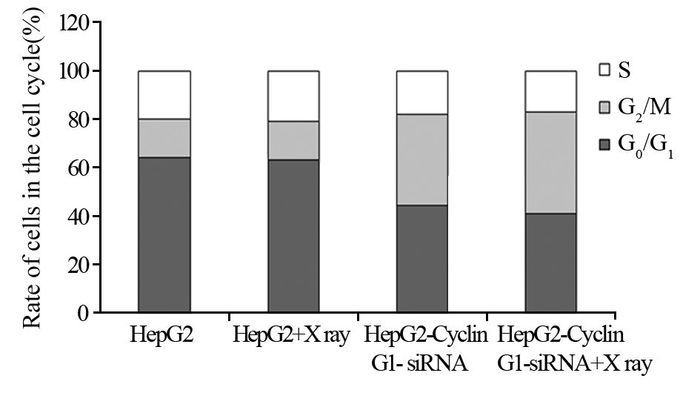
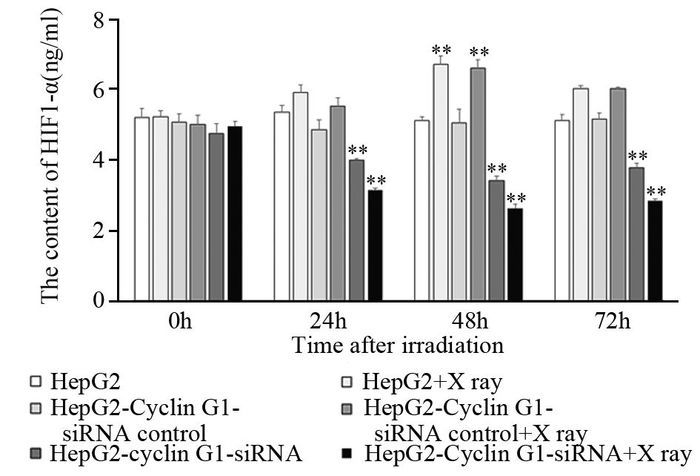
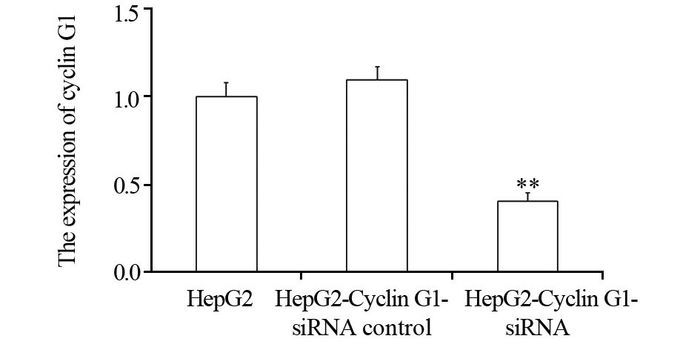
结果q-PCR显示HepG2-Cyclin G1-siRNA细胞中,Cyclin G1表达下降至正常细胞的43.5%左右,成克隆分析显示Cyclin G1-siRNA可使HepG2细胞放射敏感度增高,与正常对照组相比,放射增敏比SERDq为1.41。流式细胞分析显示HepG2-Cyclin G1-siRNA细胞与正常HepG2细胞相比,G2/M期细胞比例增加,G0/G1期细胞比例下降;ELISA显示X线照射可引起HepG2细胞的HIF-1α升高,而Cyclin G1-siRNA可以引起HIF-1α的含量显著下降。ROS含量在X射线照射后表现为轻度升高,Cyclin G1-siRNA可以使ROS含量增加,联合X线照射变化更明显。Western blot显示X线照射及Cyclin G1-siRNA均可使BCL-2表达下降及Bax表达增加,其中HepG2联合X线照射组改变最明显。
结论Cyclin G1-siRNA可以上调肝癌细胞HepG2的放射敏感度,其机制可能与调节HIF-1α、ROS及凋亡相关蛋白有关。
Abstract:ObjectiveTo investigate the effect of Cyclin G1 on the radiosensitivity of hepatocellular carcinoma HepG2 cells and its possible mechanism.
MethodsHepG2-Cyclin G1-siRNA cells was constructed through the transfection of Cyclin G1-siRNA to HepG2 cells, and the effect of Cyclin G1 on radiosensitivity of HepG2 cells was observed. Flow cytometry was used to monitor the change of cell cycle. The expression of hypoxia-inducible factor-1α(HIF-1α) and ROS were detected by ELISA analysis, and Western blot was used to detect the expression of Bcl-2 and Bax.
Resultsq-PCR showed the expression of Cyclin G1 in HepG2-Cyclin G1-siRNA cells was decreased to 43.5% of normal control group. Clone formation analysis showed that Cyclin G1-siRNA could increase the radiosensitivity of HepG2 cells, SERDq=1.41, compared with normal control group. Flow cytometry analysis showed that the cells in G2/M phase were increased and the cells in G0/G1 phase were decreased in HepG2-Cyclin G1-siRNA cells. ELISA analysis showed that the HIF-1α of HepG2 cells could be induced by X-ray irradiation, while Cyclin G1-siRNA could cause a significant decrease in the content of HIF-1α. ROS content in X-ray-irradiated cells showed mild increase, with no significant difference. Cyclin G1-siRNA could increase the content of ROS, moreover, the change was more significant when it combined with X-ray radiation. Western blot showed that X-ray irradiation and Cyclin G1-siRNA could decrease the expression of BCL-2 and increase the expression of Bax, and the changes of HepG2-Cyclin G1-siRNA combined with X-ray irradiation group were the most obvious.
ConclusionCyclin G1-siRNA could upregulate the radiosensitivity of hepatocellular carcinoma cell line HepG2, and its mechanism may be related to the regulation of HIF-1, ROS and apoptosis-related proteins expressions.
-
Key words:
- Cyclin G1 /
- Hepatocellular carcinoma /
- Radiosensitivity /
- Cell cycle /
- Hypoxia /
- Apoptosis
-
0 引言
艾滋病病毒(human immunodeficiency virus, HIV)会降低人体的免疫功能,增加人体患各种恶性肿瘤的风险[1],特别是引入联合抗反转录病毒疗法之后,艾滋病(acquired immune deficiency syndrome, AIDS)患者/HIV感染者的病程不断延长,而恶性肿瘤也逐渐成为此类患者死亡的重要病因之一[2]。近年来,我国HIV/AIDS患者人数在不断增加,HIV/AIDS合并恶性肿瘤(简称:艾滋病合并恶性肿瘤)的人数也随之增加[3-4]。我国对于艾滋病合并恶性肿瘤的规范性诊断和治疗工作开展较晚[5],加之临床上重治疗、轻随访,对于该类患者的大样本生存预后的研究报道较少。因此,本研究旨在更好地了解此类患者的临床特点,探讨患者生存预后情况及其影响因素,为进一步诊治提供参考依据。
1 资料与方法
1.1 资料
回顾性分析2013年1月1日—2020年12月31日在重庆大学附属肿瘤医院住院的354例艾滋病合并恶性肿瘤患者资料。所有患者HIV抗体检测均为阳性,均经病理确诊为恶性肿瘤,疾病编码以医院当年病案室专职编码员填写的ICD编码为依据。纳入标准:(1)首次确诊艾滋病合并恶性肿瘤患者;(2)年龄≥18岁;(3)住院治疗患者。排除标准:(1)非HIV或非恶性肿瘤患者;(2)门诊诊疗患者;(3)重复入院患者。本研究符合《赫尔辛基宣言》的要求,研究数据库已做隐私保密处理。
1.2 方法
收集患者的人口学特征(包括年龄、性别、职业、婚姻状况等)、医保信息(包括医疗保险类型、自费比例等)、住院治疗(包括确诊艾滋病合并恶性肿瘤后的住院治疗次数、平均住院天数、直接医疗总花费、是否接受手术/放疗/化疗、有无遵医嘱离院等)以及随访等预后相关信息。多因素分析采用Cox回归模型,利用条件向前法筛选变量,确定α=0.20,分类变量以哑变量的形式纳入模型,参照组为每个变量的第一个亚组,其他变量直接纳入模型进行分析。
1.3 统计学方法
采用R4.0.2,(https://www.r-project.org)统计软件进行统计分析,正态计量资料采用均数±标准差(x±s),计数资料采用绝对数(%)表示;采用Kaplan-Meier法计算平均生存时间、生存率并绘制生存曲线;单因素分析采用Log rank检验进行生存分析;多因素分析采用Cox比例风险回归模型进行分析;P < 0.05为差异有统计学意义。
2 结果
2.1 患者临床特征
患者平均年龄54.10±12.96岁(23~87岁);男女患者比例为2.1:1;所有患者中,合并淋巴瘤的患者最多,其次是肺癌和宫颈癌;从医保类型分布情况来看,城镇居民医保的患者最多。治疗数据显示:大部分患者只在医院接受一次住院治疗,确诊后在院的直接医疗费用在3万以上的有151例,占42.66%;分别接受过手术、放疗、化疗的患者占30%左右;27.97%的患者有过未遵医嘱出院的行为,见表 1。
表 1 艾滋病合并恶性肿瘤患者的临床特征Table 1 Clinical characteristics of patients with AIDS-related malignant tumor
2.2 患者生存情况
随访数据显示,54例(占15.25%)患者失访,104例(占29.38%)患者死亡,所有研究对象的平均生存时间为58.42月(95%CI: 53.11~63.73),1、3、5年观察生存率分别为78.48%、62.13%、55.31%。
单因素分析结果显示:不同恶性肿瘤类型、年龄、性别、医疗保险类型、确诊合并HIV后的住院次数、平均住院天数、直接总医疗费用、自费比例、是否放疗、有无遵医嘱离院患者的预后差异均有统计学差异,见表 2。
表 2 艾滋病合并恶性肿瘤患者预后的单因素分析Table 2 Survival of patients with AIDS-related malignant tumor: univariate analysis
2.3 影响艾滋病合并恶性肿瘤患者预后的多因素分析
多因素分析结果显示,性别、入院次数、平均住院天数、自费比例以及有无遵医嘱离院是影响患者生存预后的独立危险因素。可以认为男性患者发生死亡的风险是女性2.02倍;住院次数1次的发生死亡的风险是2次及以上的1.70倍,平均住院天数在8天及以上的患者发生死亡的风险是住院天数≤4天的2.91倍,自费比例≤40%的患者发生死亡的风险是 > 40%患者的1.52倍,遵医嘱离院的患者发生死亡的风险是未遵医嘱离院患者的2.38倍,见表 3。
表 3 影响艾滋病合并恶性肿瘤患者预后的多因素分析Table 3 Multivariate analysis of prognosis of patients with AIDS-related malignant tumor
将多因素分析有统计学差异的变量逐个绘制生存曲线图,结果显示女性患者生存预后好于男性,2次及以上的患者生存预后好于只有1次入院记录的患者,平均住院日在8天以上患者生存预后最差,自费比例 > 40%的患者生存预后逊于≤40%的患者,遵医嘱离院的患者生存预后好于未遵医嘱离院的患者,见图 1。
![]() 图 1 不同患者的Kaplan-Meier生存曲线A: survival of all and different gender patients; B: survival of patients with different admissions; C: survival of patients with different length of stay; D: survival of patients with different proportion of out-ofpocket; E: survival of patients leaving hospital according to medical advice.Figure 1 Kaplan-Meier survival curves of different groups of patients
图 1 不同患者的Kaplan-Meier生存曲线A: survival of all and different gender patients; B: survival of patients with different admissions; C: survival of patients with different length of stay; D: survival of patients with different proportion of out-ofpocket; E: survival of patients leaving hospital according to medical advice.Figure 1 Kaplan-Meier survival curves of different groups of patients3 讨论
随着我国艾滋病防控工作的进一步推进以及检测覆盖面的扩大,每年新诊断的艾滋病患者人数也在不断增加。据估计,2018年我国艾滋病新发感染人数为8万,而现存活人数达到125万,且未来几年将持续增加[6-7]。随着医疗技术的进步,艾滋病患者的生命周期会不断延长,艾滋病合并恶性肿瘤的人数也会不断增加,本次研究数据也证实了该趋势。目前国内对于艾滋病合并恶性肿瘤患者的治疗情况以及该类型患者的预后情况的研究报道不多,因此,本研究利用重庆大学附属肿瘤医院的电子病历信息系统以及随访系统,纳入HIV合并肿瘤真实世界最大病例数,回顾性地研究该类型患者的临床特点及生存预后情况。研究病例数据量较大,随访时间较长,能在一定程度上反映真实情况,为该类型患者防治工作的进一步开展提供有效依据。
艾滋病合并恶性肿瘤可以分为艾滋病相关肿瘤和非艾滋病相关肿瘤,其中艾滋病相关肿瘤主要包括卡波西肉瘤、非霍奇金淋巴瘤及浸润性宫颈癌三种级别,非艾滋病相关肿瘤主要包括肺癌、肝癌、肛门癌、皮肤癌、结直肠癌以及霍奇金淋巴瘤等[8-9]。本研究结果显示患者以艾滋病相关肿瘤为主,与国内其他研究结论一致[10-12]。具体以合并淋巴瘤最多(占28.25%),其次是肺癌(占14.97%)和宫颈癌(占14.12%),这与国内外其他研究报道有所区别[13-15],可能与不同地区的恶性肿瘤疾病构成及流行趋势不同有关,比如有些肿瘤在某些地区高发,有的医院收治的患者多,艾滋病合并该类肿瘤的患者也会相应增加。实际调查中也发现部分科室不太愿意收治艾滋病合并恶性肿瘤的患者,具体表现在门诊中发现该类患者就不收入住院治疗或者是入院发现后要求患者转院治疗。
本研究显示患者年龄以46岁及以上患者居多,性别以男性为主,与国内外研究一致[16-17],这也与恶性肿瘤发病特点一致[18]。建议临床上要加强对中老年患者艾滋病病史的询问和HIV抗体筛查。59.04%的患者只有一次入院治疗的记录、平均住院日≤4天以及直接医疗花费在1万元及以下等数据提示该类患者接受专业的抗肿瘤治疗不足。建议医院加强院感管理,设置专有病房,减少暴露风险,同时也要加强对患者的宣教和保护患者的利益。
本研究结果显示总体患者的5年观察生存率为55.31%,与国内其他研究报道的恶性肿瘤5年生存率基本一致[19]。而单艾滋病患者及时接受抗病毒治疗能够有效控制病情,5年生存率能达到98%左右[20-21]。提示艾滋病病毒只要经过系统治疗,不会影响患者抗肿瘤治疗的疗效,但具体还需要做进一步的研究来论证。多因素分析显示男性患者死亡风险高于女性,这与其他研究报道男性HIV患者死亡率高于女性结论一致[16]。可能与不同性别的患者接受抗病毒或抗肿瘤治疗的依从性或者与两性间的生活习惯不同等有关,需要进一步论证。住院次数及平均住院天数也是影响患者生存预后的因素。肿瘤患者需要定期复查,有些治疗比如放疗、化疗也是周期性的,患者需要重复住院。患者出院应该听从医嘱,如果患者没有定期住院治疗或者住院天数不够,以及患者没有遵从医嘱自行出院,可以认为患者接受治疗不足或者是治疗依从性不好,进而影响患者的预后。自费比例 > 40%的患者预后要好于≤40%的患者,提示要加大医保的投入,提高患者医疗保险比例。另外,由于本次研究对象为艾滋病合并恶性肿瘤患者的特殊人群,患者诊疗行为及预后会受到社会、经济以及医疗技术发展等多方因素影响,而本研究为单中心回顾性研究,因此建议后期联合传染病医院以及疾控中心等机构开展多中心研究。
综上所述,艾滋病合并恶性肿瘤患者的生存预后影响因素较多。只要接受系统的抗艾滋病病毒治疗,艾滋病病毒对恶性肿瘤的生存预后影响较小。因此,建议加强对患者的宣教,早发现、早系统治疗。同时也建议医院开展抗艾滋病和抗肿瘤双重治疗,做好院感防护,提高患者的生存质量。
-
![]()
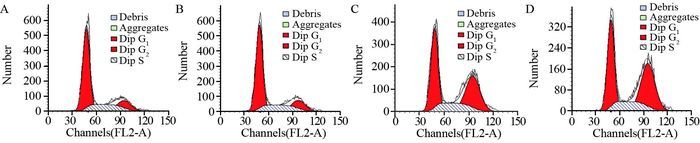
图 4 Cyclin G1对肝癌细胞HepG2细胞周期的影响
Figure 4 Effect of Cyclin G1 on cell cycle of HepG2 cells
![]()
图 6 Cyclin G1及X线照射对肝癌HepG2细胞HIF-1α表达的影响
Figure 6 Effect of Cyclin G1 and X-ray on expression of HIF-1α in HepG2 cells
![]()
图 7 CyclinG1及X线照射对肝癌细胞ROS表达的影响
Figure 7 Effect of Cyclin G1 and X-ray on expression of ROS in HepG2 cells
![]()
图 8 Cyclin G1及X线对肝癌细胞HepG2凋亡相关蛋白表达的影响
Figure 8 Effect of Cyclin G1 and X-ray on expression of apoptosis-related proteins in HepG2 cells
-
[1] Lau WY, Lai EC. Hepatocellular carcinoma: current management and recent advances[J]. Hepatobiliary Pancreat Dis Int, 2008, 7(3): 237-57. https://www.ncbi.nlm.nih.gov/pubmed/18522878
[2] Wallace MC, Preen D, Jeffrey GP, et al. The evolving epidemiology hepatocellular carcinoma: a global perspective[J]. Expert Rev Gastroenterol Hepatol, 2015, 9(6): 765-79.
[3] Schlachterman A, Craft WW Jr, Hilgenfeldt E, et al. Current and future treatments for hepatocellular carcinoma[J]. World J Gastroenterol, 2015, 21(28): 8478-91. doi: 10.3748/wjg.v21.i28.8478
[4] 于金明, 李宝生, 闫婧. 原发性肝癌的放射治疗[J]. 世界华人消化杂志, 2009, 17(10): 1005-7. doi: 10.3969/j.issn.1009-3079.2009.10.009 Yu JM, Li BS, Yan J. Radiotherapy primary hepatocellular carcinoma[J]. Shi Jie Hua Ren Xiao Hua Za Zhi, 2009, 17(10): 1005-7. doi: 10.3969/j.issn.1009-3079.2009.10.009
[5] Jensen MR, Factor VM, Fantozzi A, et al. Reduced hepatic tumor incidence in cyclin G1-deficient mice[J]. Hepatology, 2003, 37(4): 862-70. doi: 10.1053/jhep.2003.50137
[6] Wen W, Han T, Chen C, et al. Cyclin G1 expands liver tumor-initiating cells by Sox2 induction via Akt/mTOR signaling[J]. Mol Cancer Ther, 2013,12(9): 1796-804. doi: 10.1158/1535-7163.MCT-13-0099
[7] Baek WK, Kim D, Jung N, et al. Increased expression Cyclin G1 in leiomyoma compared with normal myometrium[J]. Am J Obstet Gynecol, 2003, 188(3): 634-9. doi: 10.1067/mob.2003.140
[8] Perez R, Wu N, Klipfel AA, et al. A better cell cycle target for gene therapy colorectal cancer: cyclin G[J]. J Gastrointest Surg, 2003, 7(7): 884-9. doi: 10.1007/s11605-003-0034-8
[9] Tao Y, Ruan J, Yeh SH, et al. Rapid growth a hepatocellular carcinoma and the driving mutations revealed by cell-population genetic analysis whole-genome data[J]. Proc Natl Acad Sci U S A, 2011, 108(29): 12042-7. doi: 10.1073/pnas.1108715108
[10] 滕凤猛, 吴穷, 陈昌杰. 缺氧诱导因子-1α调控肝癌细胞对奥沙利铂耐药的机制[J]. 肿瘤防治研究, 2014, 41(11): 1176-80. doi: 10.3971/j.issn.1000-8578.2014.11.004 Teng FM, WU Q, Cheng CJ. Mechanism hypoxia-inducible factor-1α inducing drug resistance to oxaliplatin in hepatic carcinoma[J]. Zhong Liu Fang Zhi Yan Jiu, 2014, 41(11): 1176-80. doi: 10.3971/j.issn.1000-8578.2014.11.004
[11] Kimura SH, Ikawa M, Ito A, et al. Cyclin G1 is involved in G2/M arrest in response to DNA damage and in growth control after damage recovery[J]. Oncogene. 2001, 20(25): 3290-300. doi: 10.1038/sj.onc.1204270
[12] Wang B, Ding YM, Fan P, et al. Expression and significance MMP2 and HIF-1α in hepatocellular carcinoma [J]. Oncol Lett, 2014, 8(2): 539-46.
[13] Yang X, Zhu H, Ge Y, et al. Melittin enhances radiosensitivity hypoxic head and neck squamous cell carcinoma by suppressing HIF-1α[J].Tumour Biol, 2014, 35(10): 10443-8. doi: 10.1007/s13277-014-2218-0
[14] Zhang YC, Jiang G, Gao H, et al. Influence ionizing radiation on ovarian carcinoma SKOV-3 xenografts in nude mice under hypoxic conditions[J]. Asian Pac J Cancer Prev, 2014, 15(5): 2353-8. doi: 10.7314/APJCP.2014.15.5.2353
[15] 张珠博, 王小春. 肿瘤细胞辐射敏感度调节新靶点—miRNAs[J]. 肿瘤防治研究, 2013, 40(12): 1193-6. doi: 10.3971/j.issn.1000-8578.2013.12.020 Zhang ZB, Wang XC. miRNAs-A new adjustable target for inradiation sensitivity tumor cells[J]. Zhong Liu Fang Zhi Yan Jiu, 2013, 40(12): 1193-6. doi: 10.3971/j.issn.1000-8578.2013.12.020
[16] 吴德华, 朱晓霞, 张洪波, 等. HIF-1α基因表达沉默对肝癌细胞的放射增敏作用[J]. 中国肿瘤临床, 2009, 36(2):110-3. http://www.cnki.com.cn/Article/CJFDTOTAL-ZGZL200902018.htm Wu DH, Zhu XX, Zhang HB, et al. Effect Silencing HIF-1α on Radiosensitization in Human Hepatocellular Carcinoma Cell Line[J]. Zhongguo Zhong Liu Lin Chuang, 2009, 36(2): 110-3. http://www.cnki.com.cn/Article/CJFDTOTAL-ZGZL200902018.htm
[17] Dayal R, Singh A, Pandey A, et al. Reactive oxygen species as mediator tumor radiosensitivity[J]. J Cancer Res Ther, 2014, 10(4): 811-8. doi: 10.4103/0973-1482.146073
[18] Sun X, Wang Q, Wang Y, et al. Brusatol Enhances the Radiosensitivity A549 Cells by Promoting ROS Production and Enhancing DNA Damage[J]. Int J Mol Sci, 2016, 17(7). pii: E997. doi: 10.3390/ijms17070997
[19] Liu S, Pan X, Yang Q, et al. MicroRNA-18a enhances the radiosensitivity cervical cancer cells by promoting radiation-induced apoptosis[J]. Oncol Rep, 2015, 33(6): 2853-62. doi: 10.3892/or.2015.3929
[20] Yang W, Sun T, Cao J, et al. Downregulation miR-210 expression inhibits proliferation, induces apoptosis and enhancesradiosensitivity in hypoxic human hepatoma cells in vitro[J]. Exp Cell Res, 2012, 318(8): 944-54. doi: 10.1016/j.yexcr.2012.02.010
-
期刊类型引用(7)
1. 张莎,唐立,闵海燕,段月勋,张娅玲,张月华,陈石芬,樊珊珊,罗佳. HIV/AIDS合并肿瘤患者生命质量状况及影响因素. 中国艾滋病性病. 2024(01): 34-38 .  百度学术
百度学术
2. 高敏. 3例艾滋病合并膀胱癌患者行手术治疗后的护理经验. 医学理论与实践. 2024(06): 1036-1037 . 百度学术
3. 亓露,李新新,郭巧丽,闫鹏,张岩红. HIV阳性女性感染高危型人乳头瘤病毒的调查分析. 临床医学. 2024(04): 54-56 . 百度学术
4. 王宁,李刚,李丹,龚胜,袁晔,周君,陈朝琼,姚晓军. 食管癌合并HIV感染/AIDS患者临床特征及淋巴结转移的危险因素分析. 肿瘤预防与治疗. 2024(04): 328-334 . 百度学术
5. 张媛媛,朱浩智,许南松,余金吨,邓创忠,黄丽华. 肿瘤专科医院2022年就诊患者常见传染病感染状况分析. 中山大学学报(医学科学版). 2024(04): 666-672 . 百度学术
6. 李新新,亓露. 艾滋病合并宫颈病变患者细胞免疫状态及其高危型HPV感染率研究. 罕少疾病杂志. 2024(09): 137-138 . 百度学术
7. 李丹,姚晓军. 食管癌合并HIV/AIDS的流行病学特征与诊治进展. 中国胸心血管外科临床杂志. 2024(11): 1684-1689 . 百度学术
其他类型引用(0)
 下载:
下载:





计量
- 文章访问数:
- HTML全文浏览量: 0
- PDF下载量:
- 被引次数: 7

