
MDM2 siRNA Inhibits Angiogenesis by Reducing Vascular Endothelial Growth Factor Production in Breast Cancer Cells
-
摘要:目的
探讨小干扰RNA(small interfering RNA, siRNA)抑制人乳腺癌细胞中鼠双微粒体2(murine double minute 2, MDM2)的表达对癌细胞中血管内皮生长因子(vascular endothelial growth factor, VEGF)合成以及裸鼠移植瘤组织内血管生成的影响。
方法根据MDM2已知的cDNA序列设计并转录合成特异性siRNA,转染高表达MDM2的人乳腺癌MDA-MB-468细胞,低氧培养后应用蛋白质印迹法和ELISA法检测肿瘤细胞及其上清液中VEGF的水平。构建裸鼠乳腺癌移植瘤模型,观察MDM2沉默后肿瘤生长情况,蛋白质印迹法、免疫组织化学及ELISA方法检测荷瘤组织和裸鼠血清标本中的VEGF含量,并以CD34标记血管内皮细胞计数微血管密度(microvessel density, MVD)。
结果siRNA抑制MDM2表达后,MDA-MB-468细胞分泌的VEGF蛋白显著减少(P=0.006)。裸鼠移植瘤模型显示,封闭MDM2表达使荷瘤组织生长变慢(P=0.008),且荷瘤小鼠血清VEGF水平明显减低(P=0.008),荷瘤组织内MVD也明显降低(P=0.003)。
结论MDM2 siRNA能有效减低乳腺癌细胞中VEGF的合成,并抑制裸鼠移植瘤组织内新生血管的生成,为MDM2/VEGF途径抗肿瘤血管生成作用的研究及靶向药物开发提供了新的思路。
Abstract:ObjectiveTo investigate the inhibitory effects of murine double minute 2 (MDM2) small interfering RNA (siRNA) on vascular endothelial growth factor(VEGF) production in human breast cancer cells and VEGF-mediated angiogenesis in xenograft tissues.
MethodssiRNA sequences of MDM2 were chemically synthesized and then transfected into human breast cancer cell line MDA-MB-468 which overexpressed MDM2. After transfection, the cells were cultured under hypoxic condition for different period of time. VEGF expression in tumor cells was measured by Western blot. The level of VEGF protein secreted in the culture supernatant was quantified by enzyme-linked immunosorbent assay (ELISA). Further, the stably-transfected cells were transplanted into the nude mice. The tumor growth was analyzed. MDM2 protein expression of tumors was determined by Western blot. Serum levels of VEGF were also measured by ELISA. VEGF expression and microvessel density (MVD) were studied by immunohistochemistry.
ResultsIn MDA-MB-468 cells, gene silencing of MDM2 resulted in the down-regulation of VEGF secretion (P=0.006) . In vivo, tumorigenicity assay showed that MDM2 depletion restrained tumor growth (P=0.008) . Serum levels of VEGF were substantially decreased (P=0.008) . MVD in transplanted tumor tissue was also significantly reduced (P=0.003) .
ConclusionMDM2 knockdown by siRNA causes a strong reduction of VEGF production in breast cancer cells and inhibition of angiogenesis in tumor transplants. The MDM2/VEGF pathway may open a new approach for anti-tumor angiogenesis and molecular targeted drugs development.
-
Key words:
- Breast neoplasms /
- RNA interference /
- MDM2 /
- VEGF
-
0 引言
肿瘤生长和转移高度依赖于瘤组织中新生血管提供丰富血供,因此抗血管生成已成为目前恶性实体肿瘤治疗的重要手段。血管内皮生长因子VEGF是迄今发现生物学活性最强的促血管生成因子[1-2]。鼠双微粒体2(murine double minute 2,MDM2)可通过转录后调控机制影响VEGF蛋白的合成,有可能成为抑制VEGF介导的肿瘤血管生成的一个新靶点。已有报道,MDM2在多数乳腺癌患者的肿瘤组织中有过度表达,并与乳腺癌的恶性生物学行为关系密切[3-4],同时发现癌组织中普遍存在的低氧环境是诱导MDM2表达增加并上调VEGF蛋白水平的重要原因[5-7]。本研究拟采取siRNA技术观察干预MDM2表达后对血管生成的抑制效果。
1 材料与方法
1.1 材料
人乳腺癌细胞系MDA-MB-468购自武汉大学中国典型培养物保藏中心;载体质粒pSUPER-neo购自美国OligoEngine公司;RPMI 1640培养液、胎牛血清、青霉素、链霉素以及RNAi-Mate转染试剂盒(货号:C-01)购自美国Hyclone公司;BALB/c-nu/nu雌性裸小鼠,4~6周龄,SPF(specificpathogenfree)条件下饲养,由华中科技大学同济医学院实验动物中心提供[实验动物合格证号:SCXK(鄂)2010-0009];MDM2抗体(鼠抗人单克隆抗体,sc-965)、鼠抗人GAPDH单克隆抗体及HRP标记的羊抗鼠、羊抗兔二抗均购自美国Santa Cruz公司;鼠抗人CD34即用型单克隆抗体购自丹麦DAKO公司;兔抗人VEGF单克隆抗体购自北京中杉公司(克隆号:SP28);VEGF ELISA试剂盒购于美国R&D公司。
1.2 方法
1.2.1 细胞培养与低氧孵育
MDA-MB-468细胞在含10%胎牛血清、50 u/ml青霉素和50 μg/ml链霉素的RPMI 1640培养液中,置于37℃、O2和CO2体积分数分别为21%和5%、恒定湿度的孵育箱内培养。细胞的低氧培养:将预处理的细胞置于O2和CO2体积分数分别为1%和5%并以氮气平衡环境中培养。
1.2.2 siRNA的体外合成与转染
根据MDM2已知的cDNA序列设计并转录合成1条含有19个核苷酸序列的siRNA(GAAGTTATTAAAGTCTGTT)(由上海生工生物工程公司合成,前期实验设计合成3条siRNA,检测各条的沉默效果,筛选找到最有效抑制MDM2蛋白表达的siRNA序列),然后将其插入pSUPER载体,得到发卡样siRNA(shRNA)表达重组质粒,同时将1条含19个核苷酸无关序列siRNA(GAGGCTATTATACTGTGAT)插入pSUPER-neo中作为阴性对照,并以未处理的pSUPER-neo作为空白对照。参照转染试剂盒说明书步骤转染和筛选,收集单细胞克隆,扩大培养,获得稳定表达pSUPER-siMDM2的MDAMB-468细胞。常氧或低氧培养3、6或12 h后分别收集细胞及其上清液。
1.2.3 乳腺癌裸鼠移植瘤模型的建立
BALB/c-nu/nu雌性裸小鼠30只,随机分为3组,每组10只:A组为MDM2 siRNA转染组;B组为无关序列转染组;C组为空白对照组。A、B、C三组接种的细胞分别为方法1.2.2中对应的三种转染细胞,以无血清细胞培养液制成5×107/ml的细胞悬液,取0.1 ml分别接种于每只裸鼠左肩胛下皮下脂肪垫。观察肿瘤出现时间,每周2次测量裸鼠瘤体大小,计算肿瘤体积,绘制生长曲线。28天后处死全部裸鼠,采血并分离血清,用于ELISA法检测VEGF水平。取皮下肿瘤称重,计算抑瘤率。肿瘤体积=长径×短径2×0.5;抑瘤率=(1-实验组瘤重/对照组瘤重)×100%。
1.2.4 蛋白质印迹法检测MDA-MB-468细胞及裸鼠荷瘤组织中MDM2和VEGF蛋白的表达
将上述从细胞和荷瘤组织中提取的总蛋白参照BCA蛋白定量试剂盒(武汉博士德生物工程有限公司)说明书进行蛋白浓度测定,分别取60 μg上样,SDS-PAGE分离蛋白,电转移至硝酸纤维素膜(武汉博士德生物工程有限公司提供)上。5%脱脂牛奶室温封闭1 h后,加入一抗(MDM2 1:200,VEGF 1:200,GAPDH 1:500)4℃孵育过夜。TBST洗膜3次,每次15 min,接着加入HRP标记二抗(1:10 000)室温孵育1 h,然后TBST洗膜3次,每次15 min。最后使用ECL化学发光法(武汉博士德生物工程有限公司)进行显影。
1.2.5 ELISA检测MDA-MB-468细胞上清液及裸鼠血清中VEGF的含量
转染后的3组细胞分别置于常氧和低氧环境中培养3、6或12 h,收集细胞上清液,参照ELISA试剂盒说明书检测细胞上清液中VEGF的含量。另外,取转染后各组肿瘤细胞接种第28天的小鼠,从其眼眶后静脉丛采血,并分离血清,ELISA检测各组裸鼠血清标本中VEGF的浓度。
1.2.6 免疫组织化学检测荷瘤组织VEGF蛋白表达及微血管密度(microvessel density,MVD)
荷瘤组织甲醛固定后,制作石蜡切片,脱蜡修复后分别用VEGF单抗或CD34单抗室温孵育1 h,冲洗后用Envision二抗(DAKO公司)孵育0.5 h,DAB显色,苏木精对比染色,CD34标记组织内血管内皮细胞,低倍镜(×40)下确定3个血管着色最密集的区域,高倍镜(×200)下计数微血管数目,取10个视野的均值作为MVD。
1.3 统计学方法
采用SPSS19.0统计软件进行数据分析。每组实验均重复3次,各实验结果以均数±标准差表示,多组样本均数比较进行方差齐性检验,组间比较采用单因素方差分析(One-way ANOVA),方差不齐者采用非参数检验(Kruskal-Wallistest)。以P<0.05为差异有统计学意义。
2 结果
2.1 MDM2 siRNA转染细胞和荷瘤组织中MDM2和VEGF蛋白的表达情况
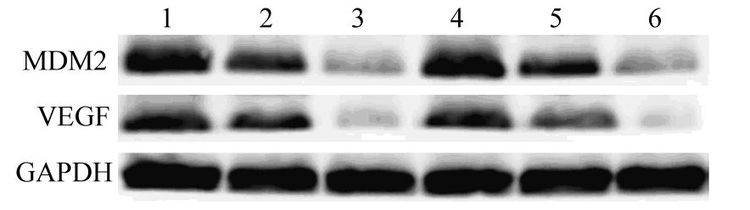
与空白对照组和无关序列转染组相比,MDM2siRNA转染组细胞MDM2蛋白表达水平显著下调(分别P=0.003和P=0.004),提示MDM2蛋白的表达被有效抑制,而无关序列转染组和空白对照组比较差异无统计学意义(P=0.890),见图 1。同样,MDM2蛋白的沉默效果在裸鼠荷瘤组织中也得以证实。与空白对照组和无关序列转染组相比,MDM2 siRNA转染组肿瘤组织MDM2蛋白表达水平显著减低(分别P=0.009和P=0.009),而无关序列转染组和空白对照组比较差异无统计学意义(P=0.823)。在上述分组检测MDM2的同时也对VEGF蛋白水平进行了检测,观察到与MDM2类似的变化。MDM2 siRNA转染后,无论是肿瘤细胞还是裸鼠荷瘤组织中VEGF的表达均显著减低,明显低于空白对照组和无关序列转染组(肿瘤细胞:分别P=0.007和P=0.006;裸鼠荷瘤组织:分别P=0.008和P=0.006),而无关序列转染组和空白对照组比较差异无统计学意义(肿瘤细胞:P=0.781;裸鼠荷瘤组织:P=0.745)。
![]() 图 1 蛋白质印迹法检测MDM2 siRNA转染细胞和荷瘤组织中MDM2和VEGF蛋白的表达情况Figure 1 Expressions of MDM2 and VEGF in MDAMB-468 cells and xenografts determined by Western blot1-3: MDA-MB-468 cells; 4-6: xenografts; 1 and 4: blank control groups;2 and 5: siRNA control groups; 3 and 6: MDM2 siRNA groups; MDM2:murine double minute 2; VEGF: vascular endothelial growth factor
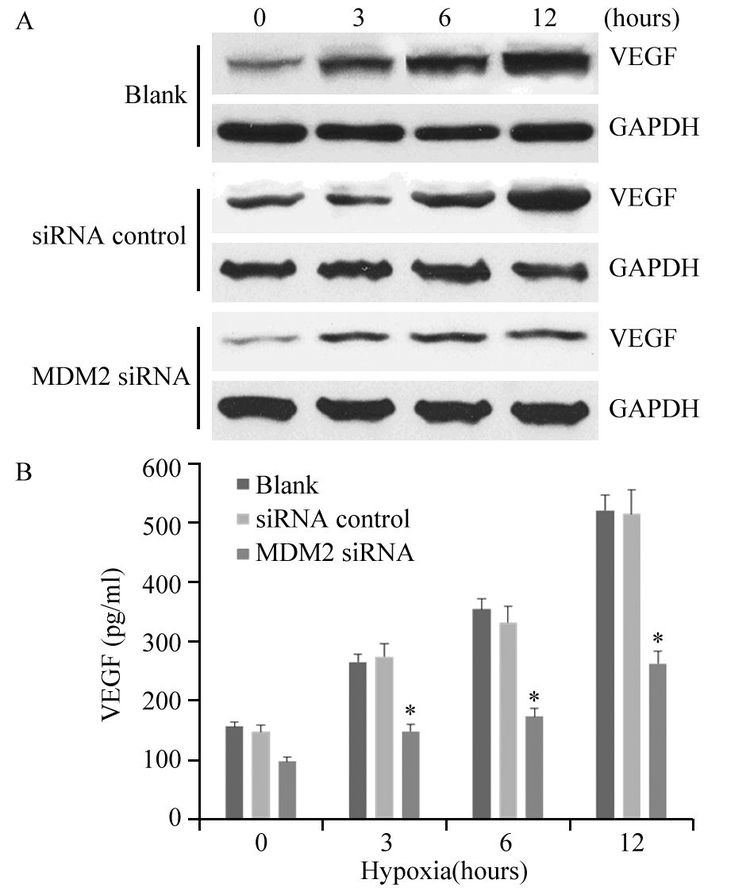
图 1 蛋白质印迹法检测MDM2 siRNA转染细胞和荷瘤组织中MDM2和VEGF蛋白的表达情况Figure 1 Expressions of MDM2 and VEGF in MDAMB-468 cells and xenografts determined by Western blot1-3: MDA-MB-468 cells; 4-6: xenografts; 1 and 4: blank control groups;2 and 5: siRNA control groups; 3 and 6: MDM2 siRNA groups; MDM2:murine double minute 2; VEGF: vascular endothelial growth factor2.2 MDM2 siRNA抑制缺氧环境中肿瘤细胞VEGF的生成
图 2显示缺氧环境能够刺激肿瘤细胞VEGF的生成,并且随缺氧时间延长,MDA-MB-468细胞生成的VEGF水平逐渐增高。然而,MDM2干扰后VEGF的生成受抑。Western blot结果显示各缺氧时间点MDM2 siRNA转染组细胞内VEGF蛋白的含量均明显低于空白对照组(3 h: P=0.037;6 h: P=0.022;12 h: P=0.004)和无关序列转染组(3 h: P=0.046;6 h: P=0.023;12 h: P=0.003)。ELISA检测结果显示各缺氧时间点MDM2 siRNA转染组细胞培养上清液中VEGF的含量均明显低于空白对照组(3 h: P=0.043;6 h: P=0.011;12 h:P=0.006)和无关序列转染组(3 h: P=0.040;6 h:P=0.036;12 h: P=0.004)。
![]() 图 2 MDM2 siRNA抑制缺氧环境中肿瘤细胞内(A)和培养上清液中(B)VEGF的水平Figure 2 Inhibitory effects of MDM2 siRNA on hypoxiainduced VEGF production in tumor cells(A) and culture supernatant(B)A: VEGF expressions in the tumor cells were determined by Western blot; B: the concentrations of VEGF in the culture supernatant were determined by ELISA; *: P<0.01, compared with blank and siRNA control groups
图 2 MDM2 siRNA抑制缺氧环境中肿瘤细胞内(A)和培养上清液中(B)VEGF的水平Figure 2 Inhibitory effects of MDM2 siRNA on hypoxiainduced VEGF production in tumor cells(A) and culture supernatant(B)A: VEGF expressions in the tumor cells were determined by Western blot; B: the concentrations of VEGF in the culture supernatant were determined by ELISA; *: P<0.01, compared with blank and siRNA control groups2.3 MDM2 siRNA抑制裸鼠体内VEGF及荷瘤组织内新生血管的生成
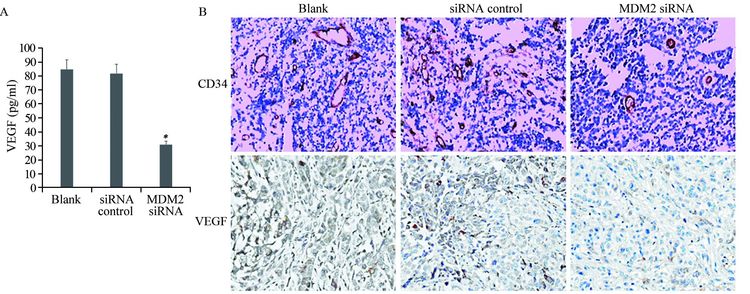
ELISA检测结果显示,MDM2 siRNA转染组血清VEGF水平明显低于空白对照组和无关序列转染组(分别P=0.008和P=0.005),而无关序列转染组和空白对照组之间差异无统计学意义(P=0.860),见图 3A。免疫组织化学法检测原位肿瘤组织中VEGF的表达,并以CD34染色计数微血管生成,结果显示MDM2沉默后VEGF蛋白表达出现下降,而空白对照组、无关序列转染组和MDM2 siRNA转染组的MVD分别为(41.07±7.59)、(39.46±6.77)和(21.25±4.07),提示siRNA阻断MDM2的表达显著抑制了肿瘤组织内新生血管的生成(P=0.003),见图 3B。
![]() 图 3 MDM2 siRNA抑制裸鼠体内VEGF及荷瘤组织内新生血管的生成Figure 3 Inhibitory effect of MDM2 siRNA on VEGF-mediated angiogenesisA: the concentrations of VEGF in the serum sample of nude mice were determined by ELISA; *: P<0.01, compared with blank and siRNA control groups; B: representative histological comparisons of VEGF production and tumor vessel content in xenografts from different groups, as stained by VEGF and CD34 immunohistochemistry (Envision ×200) . MVD was lower in MDM siRNA group than in blank and siRNA control groups
图 3 MDM2 siRNA抑制裸鼠体内VEGF及荷瘤组织内新生血管的生成Figure 3 Inhibitory effect of MDM2 siRNA on VEGF-mediated angiogenesisA: the concentrations of VEGF in the serum sample of nude mice were determined by ELISA; *: P<0.01, compared with blank and siRNA control groups; B: representative histological comparisons of VEGF production and tumor vessel content in xenografts from different groups, as stained by VEGF and CD34 immunohistochemistry (Envision ×200) . MVD was lower in MDM siRNA group than in blank and siRNA control groups2.4 MDM2 siRNA对裸鼠移植瘤的生长有抑制作用
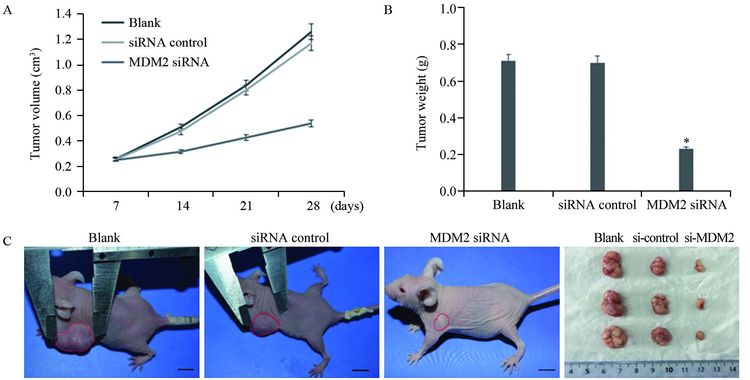
转染后细胞接种裸鼠28天后测量移植瘤体积,空白对照组、无关序列转染组和MDM2 siRNA转染组分别为(1.26±0.37)cm3、(1.17±0.29)cm3和(0.52±0.18)cm3;瘤重分别为(0.71±0.17)g、(0.70±0.14)g和(0.23±0.09)g,MDM2 siRNA转染组移植瘤体积(P=0.009和P=0.008)和瘤重(P=0.008和P=0.007)均显著低于空白对照组和无关序列转染组,抑瘤率达到69%,见图 4。
![]() 图 4 MDM2 siRNA对裸鼠移植瘤的生长的抑制作用Figure 4 Inhibitory effect of MDM siRNA on tumor growthA: xenograft tumor size; B: xenograft tumor weight; C: representative pictures from each group. *: P<0.01, compared with blank and siRNA control groups
图 4 MDM2 siRNA对裸鼠移植瘤的生长的抑制作用Figure 4 Inhibitory effect of MDM siRNA on tumor growthA: xenograft tumor size; B: xenograft tumor weight; C: representative pictures from each group. *: P<0.01, compared with blank and siRNA control groups3 讨论
MDM2作为p53的经典抑制因子为人们所熟知,它通过阻断p53介导的基因转录和降解p53来实现其癌蛋白功能[8]。对MDM2的研究已广泛而深入,但多局限于癌细胞凋亡、抗增殖以及细胞周期停滞等方面[9-10]。然而,逐渐有越来越多的证据显示MDM2可以不依赖于p53发挥其癌蛋白的功能。无野生型p53而MDM2表达增高的肿瘤细胞仍然具有高侵袭潜能,且患者往往出现化疗失败的情况[11]。本研究关注于MDM2在VEGF所介导的肿瘤血管生成中的非p53依赖调控机制。我们在前期研究中已初步证实缺氧状态可诱导肿瘤细胞内MDM2蛋白从细胞核向细胞质转位,细胞质内MDM2蛋白其COOH末端的环状结构可直接结合于VEGF mRNA的 3’非翻译区,这种结合将增加VEGF mRNA的稳定性,进而促进VEGF蛋白的翻译合成[6]。
VEGF诱导的血管生成对实体瘤生长发挥关键作用,且与肿瘤转移关系密切[12]。肿瘤组织内VEGF的表达受多种因素影响,其中低氧诱导的VEGF过度表达是最令人关注的[13]。实体肿瘤组织在低氧状态时,血供不足以及组织中代谢产物的减少均可诱导VEGF表达增加,导致代偿性新生血管生成,从而满足肿瘤生长的需要。研究表明低氧可以在转录和转录后水平同时影响VEGF的表达,如广为人知的缺氧诱导因子1α(hypoxia-induciblefactor 1α,HIF-1α)就是通过与VEGF启动子结合在转录水平调节VEGF的表达[14]。为探索VEGF可能存在的其他调控途径,本实验模拟低氧环境并用siRNA阻断乳腺癌细胞内MDM2蛋白的合成来观察对VEGF生成的影响,发现MDM2的去表达可使乳腺癌细胞分泌的VEGF蛋白显著减少。除了在体外模型中发现下调乳腺癌细胞中MDM2的表达可有效抑制肿瘤细胞合成VEGF外,我们在体内模型中也观察到类似效应,敲除MDM2的移植瘤在裸鼠体内生长变慢,荷瘤小鼠血清VEGF水平也出现降低,荷瘤组织内VEGF蛋白表达也有所降低且新生血管明显减少。这些现象显示MDM2对VEGF的合成具有调控作用,从而影响肿瘤组织血管生成。抑制MDM2可解除对p53的拮抗,恢复p53促凋亡活性,也可以同时通过不依赖于p53的途径减弱其他生长因子或癌蛋白的表达,而正是这些因素共同的作用才构成肿瘤发生发展的复杂性。
我们前期已使用ASO和nutlin-3两种作用机制不同的分子初步验证了MDM2对VEGF的正向调控效应,且该效应并不依赖p53,nutlin-3干扰MDM2与p53的结合,但不抑制VEGF的表达;而ASO则是直接阻断MDM2 mRNA,阻碍其蛋白合成,从而影响VEGF水平[7]。本文在此理论基础上,继续研究该调控机制在肿瘤靶向治疗中的潜在价值,以p53突变的MDA-MB-468为对象,采用其他不同于ASO的反义技术如siRNA干预MDM2合成来观察其抗血管生成效应,比较各种反义技术的效果,以期提供更多的选择。siRNA和ASO原理和技术方法相近,且需要进行化学修饰来保护其不被核酸酶迅速降解,这在一定程度上削弱了其导入细胞的效率,相比之下siRNA具有更强的优势和潜力。
虽然可利用反义寡核苷酸有效靶位点设计RNA干扰序列,但我们在预实验中发现采用前文ASO序列(5’-UGACACCTGTTCTCACUCAC-3’)来设计相对应的siRNA,其对MDM2的抑制效果并不显著。本实验采用另外一段不同的siRNA序列却获得了理想的阻断效果,推测可能是由于结合位点或者是mRNA二级结构的不同导致了这些差异,这提示靶位点的选择可能是反义技术设计中的一个关键点。并且我们还观察到ASO处理的细胞起效较慢,约12 h后才出现显著的抑制效果,但持续时间较长,而用siRNA处理的细胞3 h就出现VEGF水平下降,至12 h达到抑制峰值。虽然两个实验的细胞处理方式有所不同,但也提示siRNA起效时间较早、持续时间较短的特点,这在其他报道中也已经证实[15]。这可能与ASO经过化学修饰,细胞穿透性较弱,但稳定性比siRNA高有关。因此,ASO和siRNA干扰在实验中都取得了确切的结果,这两种反义技术各有长处,互为补充,将为以后设计药物合成路径及应用方案提供更多选择。
综上,本研究系列实验结果给我们开辟了一条新的思路,即不单纯采用bevacizumab或VEGF受体负显性突变子来抑制肿瘤组织内VEGF的过量分泌,而是寻找新的靶点更有效地达到这个目的。那么,MDM2是其中一个有价值的选择,尤其是在MDM2过度表达的肿瘤组织中,通过各种反义技术的靶向干预减少它的产生将有助于抑制肿瘤组织中VEGF的过度表达。
-
![]()
图 1 蛋白质印迹法检测MDM2 siRNA转染细胞和荷瘤组织中MDM2和VEGF蛋白的表达情况
Figure 1 Expressions of MDM2 and VEGF in MDAMB-468 cells and xenografts determined by Western blot
![]()
图 2 MDM2 siRNA抑制缺氧环境中肿瘤细胞内(A)和培养上清液中(B)VEGF的水平
Figure 2 Inhibitory effects of MDM2 siRNA on hypoxiainduced VEGF production in tumor cells(A) and culture supernatant(B)
![]()
图 3 MDM2 siRNA抑制裸鼠体内VEGF及荷瘤组织内新生血管的生成
Figure 3 Inhibitory effect of MDM2 siRNA on VEGF-mediated angiogenesis
-
[1] Bassal R, Schejter E, Bachar R, et al. Recent trends of cervical cancer and Cervical Intraepithelial Neoplasia 3(CIN3) in Israel[J]. Arch Gynecol Obstet, 2015, 292(2) : 405-13.
[1] Schneider BP, Gray RJ, Radovich M, et al. Prognostic and predictive value of tumor vascular endothelial growth factor gene amplification in metastatic breast cancer treated with paclitaxel with and without bevacizumab; results from ECOG 2100 trial[J]. Clin Cancer Res, 2013, 19(5): 1281-9. [2] Croci DO, Cerliani JP, Dalotto-Moreno T, et al. Glycosylationdependent lectin-receptor interactions preserve angiogenesis in anti-VEGF refractory tumors[J]. Cell, 2014, 156(4): 744-58. [2] Touboul C, Skalli D, Guillo E, et al. Treatment of cervical cancer[J]. Rev Prat, 2014, 64(6) : 802-6.
[3] Sato I, Umemura M, Mitsudo K, et al. Hyperthermia generated with ferucarbotran (Resovist(A (R))) in an alternating magnetic field enhances cisplatin-induced apoptosis of cultured human oral cancer cells[J]. J Physiol Sci, 2014, 64(3) : 177-83.
[3] Maguire M, Nield PC, Devling T, et al. MDM2 regulates dihydrofolate reductase activity through monoubiquitination[J]. Cancer Res, 2008, 68(9): 3232-42. [4] Shi H, Liu T, Fu C, et al. Insights into a microwave susceptible agent for minimally invasive microwave tumor thermal therapy[J]. Biomaterials, 2015, 44: 91-102.
[4] Wang W, Qin JJ, Voruganti S, et al. The pyrido[b]indole MDM2 inhibitor SP-141 exerts potent therapeutic effects in breast cancer models[J]. Nat Commun, 2014, 5: 5086. [5] Tropea A, Biondi A, Corsaro A, et al. Combined microwave thermal ablation and liver resection for single step treatment of otherwise unresectable colorectal liver metastases; a monoistitutional experiences[J]. Eur Rev Med Pharmacol Sci, 2014, 18(2 Suppl): 6-10.
[5] Carroll VA, Ashcroft M. Regulation of angiogenic factors by HDM2 in renal cell carcinoma[J]. Cancer Res, 2008, 68(2): 545-52. [6] Zhou S, Gu L, He J, et al. MDM2 regulates vascular endothelial growth factor mRNA stabilization in hypoxia[J]. Mol Cell Biol, 20 11, 31(24): 4928-37. [6] Sun F, Yin LF, Zhou JP. Current advances in study of thermosensitive liposomes[J]. Yao Xue Jin Zhan, 2010, 34(9) : 399-405.
[7] Xiong J, Yang Q, Li J, et al. Effects of MDM2 inhibitors on vascular endothelial growth factor-mediated tumor angiogenesis in human breast cancer[J]. Angiogenesis, 2014, 17(1): 37-50. [7] 孙飞, 尹莉芳, 周建平. 热敏脂质体的研究进展[J]. 药学进展, 2010, 34(9) : 399-405.
[8] Mohammad F, Yusof NA. Doxorubicin-loaded magnetic gold nanoshells for a combination therapy of hyperthermia and drug delivery[J]. J Colloid Interface Sci, 2014, 434: 89-97.
[8] Gajjar M, Candeias MM, Malbert-Colas L, et al. The p53 mRNAMdm2 interaction controls Mdm2 nuclear trafficking and is required for p53 activation following DNA damage[J]. Cancer Cell, 2012, 21(1): 25-35. [9] Chen YT, Yao HR, Xu LF, et al. Hepatic arterial infusion chemotherapy combined with endogenetic hyperthermia treatment of hilar cholangiocarcinoma[J]. Hepatogastroenterology, 2014, 61(129) : 151-5.
[9] Huang M, Zhang H, Liu T, et al. Triptolide inhibits MDM2 and induces apoptosis in acute lymphoblastic leukemia cells through a p53-independent pathway[J]. Mol Cancer Ther, 2013, 12(2): 18 4-94. [10] Yang TT, Wu JF, Li XJ, et al. Effect of Gene Mutation of p53 in Lung Cancer on TSG101/MDM2 Signal Passway[J].Zhong Liu Fang Zhi Yan Jiu, 2011, 38(7): 774-7. [杨廷桐, 武俊芳, 李秀杰, 等. p53基因突变对非小细胞肺癌TSG101/MDM2信号通路的 影响[J] 肿瘤防治研究, 2011, 38(7): 774-7.] [10] Zhang Z, Wang J, Nie X, et al. Near infrared laser-induced targeted cancer therapy using thermoresponsive polymer encapsulated gold nanorods[J]. J Am Cheml Soc, 2014, 136(20) : 7317-26. Lee SM, Kim HJ, Kim SY, et al. Drug-loaded gold plasmonic nanoparticles for treatment of multidrug resistance in cancer[J]. Biomaterials, 2014, 35(7) : 2272-82.
[11] Nag S, Zhang X, Srivenugopal KS, et al. Targeting MDM2-p53 interaction for cancer therapy: are we there yet?[J]. Curr Med Chem, 2014, 21(5): 553-74. [11] You J, Zhang P, Hu F, et al. Near-infrared light-sensitive liposomes for the enhanced photothermal tumor treatment by the combination with chemotherapy[J]. Pharm Res, 2014, 31(3) : 554-65.
[12] Kouloulias V, Triantopoulou S, Vrouvas J, et al. Combined chemoradiotherapy with local microwave hyperthermia for treatment of T3N0 laryngeal carcinoma: a retrospective study with long-term follow-up[J]. Acta Otorhinolaryngol Ital, 2014, 34(3) : 167-73.
[12] Di Benedetto M, Toullec A, Buteau-Lozano H, et al. MDAMB- 23 1 breast cancer cells overexpressing single VEGF isoforms display distinct colonisation characteristics[J]. Br J Cancer, 2015, 11 3(5): 773-85. [13] Mésange P, Poindessous V, Sabbah M, et al. Intrinsic bevacizumab resistance is associated with prolonged activation of autocrine VEGF signaling and hypoxia tolerance in colorectal cancer cells and can be overcome by nintedanib, a small molecule angiokinase inhibitor[J]. Oncotarget, 2014, 5(13): 4709-21. [13] Zhou Y, Han G, Wang Y, et al. Radiofrequency heat-enhanced chemotherapy for breast cancer: towards interventional molecular image-guided chemotherapy[J]. Theranostics, 2014, 4(11) : 1145-52.
[14] Pereira ER, Frudd K, Awad W, et al. Endoplasmic reticulum (ER) stress and hypoxia response pathways interact to potentiate hypoxia-inducible factor 1 (HIF-1) transcriptional activity on targets like vascular endothelial growth factor (VEGF)[J]. J Biol Chem, 2014, 289(6): 3352-64. [14] DeWitt MR, Pekkanen AM, Robertson J, et al. Influence of hyperthermia on efficacy and uptake of carbon nanohorn-cisplatin conjugates[J]. J Biomech Eng, 2014, 136(2) : 021003.
[15] Chen WY, Chen Y, Yang R, et al. The preparation of doxorubicinloaded Fe3O4 magnetic nanoparticles modified with PEG and the effect of SKOV-3 cells[J]. Zhongguo Shi Yan Zhen Duan Xue, 2014, 18(6) : 880-3. Wang L, Shi J, Jia X, et al. NIR-/pH-Responsive drug delivery of functionalized single-walled carbon nanotubes for potential application in cancer chemo-photothermal therapy[J]. Pharm Res, 2013, 30(11) : 2757-71.
[15] Li RW, Zhao YY, Yang ZC, et al. siRNA Inhibits MDM2 Expression and Cell Proliferation in PC-3 Cell Line[J]. Zhong Liu Fang Zhi Yan Jiu, 2009, 36(1): 24-7. [李然伟, 赵燕颖, 杨泽城, 等. siRNA抑制人前列腺癌细胞PC3的MDM2表达及细胞增殖[J]. 肿瘤防治研究, 2009, 36(1): 24-7.]
 下载:
下载:
计量
- 文章访问数: 1182
- HTML全文浏览量: 344
- PDF下载量: 338

